![[Laboratory VII - The Origin of Seed Plants]](OspD/OspfontN.jpeg)
The earliest known seed plants are reported from the Late Devonian (Famennian) of West Virginia (Rothwell et al., 1989). Slender branch systems bear incompletely-fused cupules containing four upright ovules. In each ovule, the nucellus -- megasporangium -- is almost completely surrounded by an integument that developed from four fused tissue lobes. Each lobe is vascularized by a trace from the sporophyte axis. A slightly younger Late Devonian seed, Archaeosperma, has a similar loose arrangement of cupule and integument, which appears to be derived from the ultimate branching system of a plant like Aneurophyton. Unfortunately, we have little information about the vegetative structures associated with these early ovules. Consequently, their link to the progymnosperms remains hazy.
To find a seed plant for which vegetative and reproductive structures are well known, we have to jump forward in time to the early Pennsylvanian. The stem genus Lyginopteris is characterized by a eustele, bifacial cambium, manoxylic wood, and a well developed cortex characterized by broad sclerenchyma fibers in an anastomosing network (VG 1:8). Manoxylic wood is one feature that links some of the early seed plants with the cycads and related lineages, which we will study next week. However, there is no clear differentiation between manoxylic wood, which has abundant parenchyma scattered between tracheids, and pycnoxylic wood, which is parenchyma poor. Instead, the amount of parenchyma varies from groups to group in a continuous range of variation.
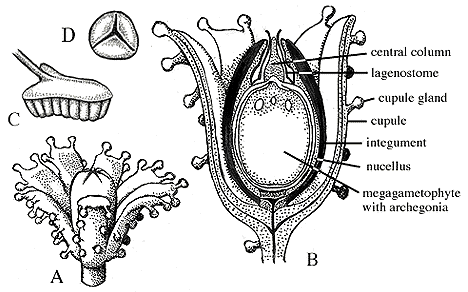
Lyginopteris has axillary branching and a small petiole with a V- or W-shaped leaf trace. This feature suggests that the leaves of Lyginopteris are developmentally distinct from the plant's branch system. Fronds commonly have a branched rachis that bore Sphenopteris (VG 1:9) or Pecopteris foliage. Note that Pecopteris is familiar from the Carboniferous Marattialian tree ferns (e.g., Psaronius). Ovules, called Lagenostoma, are small, circular in cross section (radiospermic) (VG 1:10), and borne in cup-like structures of sterile tissue (cupules) (Figure 7.8). The apex of the megasporangium contained within the ovule is elaborated into a structure to help direct pollen to the fertile areas. Pollen organs -- Crossotheca -- are small and displayed in clusters. Pollen is trilete (Figure 7.8).
|
| Figure 7.8: Organs associated with Lyginopteris stems. (A) Ovule and cupule with distinctive glands that allowed connection of these structures with Lyginopteris stems and associated organs. (B) Longitudinal section of ovule Lagenostoma. (C) Possible reconstruction of pollen organ, Crossotheca, showing pollen sacs pendant from a pad of tissue. (D) Pollen produced by Crossotheca. Note the plesiomorphic trilete mark. Pollen with this plesiomorphic aperture are sometimes called "prepollen".
|
You will recall that the trilete morphology is characteristic of spores produced by meiosis of a zygote or other diploid spore mother cell. Why, then, are we calling these grains pollen? Pollen is generally defined as the microgametophyte and its associated protective shell in ovule-producing (e.g., seed) plants. Consequently, we call the trilete pollen of Lyginopteris "pollen" because it is functioning as pollen in a plant that produces an ovule. The morphology doesn't catch up until a bit later. Some workers have coined the term "prepollen" to refer to plesiomorphic spore morphologies that function as pollen grains.
A variety of other ovules are reported from the latest Devonian, Mississippian and early Pennsylvanian. Many have unusual morphology and together they suggest that the seed plant lineage was experimenting with many new forms. Some workers have suggested that extensive morphological experimentation early in the history of lineages occurs because the new organisms are moving into new ecological "spaces" where they are suffering little competition. This allows many morphologies to coexist. Later, when members of the new lineage begin to compete with one another, many of the early morphologies are removed by selection. Others speculate that morphological variety early in lineages is the result of poorly-canalized developmental pathways. Regardless of cause, by the middle of the Pennsylvanian, the experimental phase is largely over and a few basic designs become common. At this point, seed plants begin to become more species rich.
|
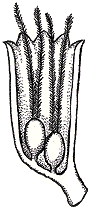
Figure 7.9: Gnetopsis from the Pennsylvanian showing two ovules in a single cupule and modified, feather-like extensions of the micropyle that may have aided in pollen capture (VG 1:11). |
![[Previous Page]](../VPLimg/Back.jpeg) |
![[Title Page]](OspD/Ospbutt.jpeg) |
![[Glossary]](../VPLimg/Glossbutt.jpeg) |
![[Range Chart]](../VPLimg/Rangebutt.jpeg) |
![[Geologic Time Scale]](../VPLimg/timesbutt.jpeg) |
![[Next Page]](../VPLimg/Forward.jpeg) |


{kind=link}
{kind=link}
{kind=link}
{kind=link}