![[Laboratory X - Anthophytes, Glossopterids and Others]](CupD/CupfontB.jpeg)
If we are to look for the earliest true angiosperms, we must first know what separates the flowering plants from the other seed plants. The obvious answer is flowers! However, other Mesozoic seed plants came perilously close to the "flower habit" without actually crossing the biological line to angiosperm-ness. A variety of characters have been proposed to distinguish angiosperms. Some of those that might be recognized in the fossil record are summarized here:

|
Figure 10.11: Tectate-collumellate wall (exine) typical of angiosperm pollen. |
Despite all of this interest and introspection, the origin and ancestry of the flowering plants remains a mystery -- though perhaps not an abominable one. No clear ancestral lineage has been identified beyond the complex of Mesozoic seed plants from which all of the major lineages branched. Attempts to speculate on angiosperm ancestors using Early Cretaceous fossils has been complicated by the mis-assignment of many early angiosperm fossils to modern or nearly modern taxa (families and genera). Research taking a phylogenetic approach using modern angiosperms to speculate on flowering-plant origins has been thwarted by the antiquity and apparently rapid diversification of the group -- two factors that tend to blur relationships. Phylogenetic studies that have attempted to unite fossil and modern taxa have probably yielded the best results to date, but these are complicated by basic questions of homology between the flowering plants and the strange seed plants of the Mesozoic. Sound familiar?
Despite what we still do not know, much can be said about the earliest flowering plants. (1) The first true angiosperms appear in the fossil record in the Early Cretaceous. Some have suggested more ancient roots for this group based on its early diversity. In the scenario, the early angiosperms lived in upland, possibly arid, regions where they were unlikely to enter the fossil record. The lack of any trace (including pollen, which can be widely transported) of these pre-Cretaceous angiosperms makes this supposition increasingly unlikely, particularly as more previously-unexplored regions become known. (2) The cradle of the flowering plants was probably the Cretaceous tropics, particularly the northern portion of the Gondwanan continent. This hypothesis is based on the observation that the oldest records of angiosperm remains (primarily pollen, which can be definitively recognized) are found at low paleolatitudes and spread north and south through time. (3) The earliest angiosperms have a mix of features that, in the modern flora, are diagnostic of major groups (families or higher). This was a puzzle that led to much mis-classification of early angiosperms. However, the observation probably should be telling us something about the patterns of morphologic change in the early stages of diversification of a new clade.
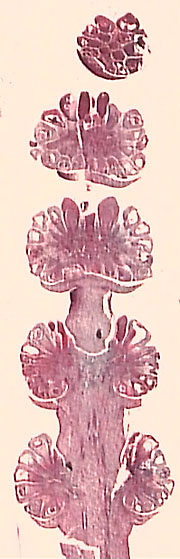
Among the candidates for pre-Cretaceous angiosperms is Sanmiguelia lewisii (Figure 10.12) (VG 3:1) from the Upper Triassic of Colorado. The foliage of this controversial plant is reminiscent of palms and several other monocot groups in overall aspect (parallel veins and clasping petioles). However, these characteristics are not necessarily those phylogenetic analysis predicts for the most primitive monocots. Consequently, the relationship of Sanmiguelia to the monocots -- or even the angiosperms -- remains in question.

|
| Figure 10.12: Sanmiguelia lewisii. (A) Axelrodia, the cupulate, ovuliferous "inflorescence" emerging from leaf axils. (B) Pollen-bearing "inflorescence", Synangispadixis.
|
Some reproductive structures have been found in association with Sanmiguelia. However, their relationship with the foliage is not clear and they are not well preserved. Consequently, they have not clarified the relationship of this plant to the angiosperms. If it is an angiosperm at all, Sanmiguelia, would likely lay near the phylogenetic split between the monocots and dicots.
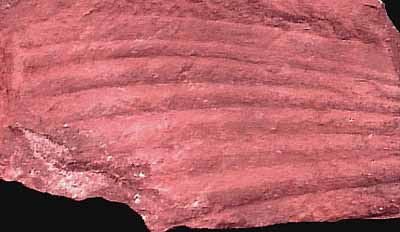
Second only to pollen, leaves are the most common fossils of early flowering plants. As such, they have become one of the important keys to understanding patterns of angiosperm diversification during the Cretaceous. Detailed studies of over-all shape, venation, and tooth pattern have demonstrated an increase in diversity, complexity and organization through time (Figure 10.13) (VG 3:2)(VG 3:3). Early angiosperm leaves, for example, have relatively disorganized patterns of vein branching, while later representatives have a well-developed hierarchy of veins that form regular patterns that are useful for recognizing taxonomic groups.

|
| Figure 10.13: A variety of Early Cretaceous leaves from the Potomac Group from eastern North America.
|
Early work on fossil angiosperm leaves focused on a picture-matching approach to allying fossil forms to modern families or genera. This approach led to the erroneous impression that many of the extant families of flowering plants were present and well-differentiated by the middle part of the Cretaceous. With the discovery of more fossils, particularly those of flowers, angiosperm paleontologists began to realize that the earliest angiosperms were commonly a mix of morphological features that today define extant families. This realization forced a reevaluation of the earlier work on dicot leaves. Work today focuses on detailed analytical study of morphology, venation pattern, and cellular structure of cuticle, which has led to important insights into the patterns of the early radiation of flowering plants.
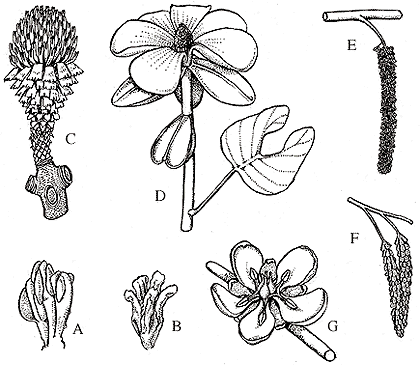
As you might imagine, flowers are among the rarest of plant fossils. Because these structures are both ephemeral and delicate, extraordinary circumstances are required to preserve them in the rock record. Nonetheless, there is a substantial body of descriptions of Cretaceous and early Tertiary flowers (for example platanecous flowers from the Clarno Flora (VG 3:4))some of which is listed in the "Read More About It" bibliography at the end of the lab. The rarity of fossil flowers has been a frustration for angiosperm paleobotanists because flowers (reproductive structures in general) are among the most important features for classifying and recognizing relationships among flowering-plant groups. Consequently, each new fossil find takes on tremendous importance and commonly re-arranges our notions about the basal relationships of the flowering plants. Unfortunately, our teaching collection does not include any fossil flowers, but Figure 10.14 illustrates reconstructions of some of the variety already present in the Cretaceous.
Fruits and seeds, particularly those with hard, protective shells are somewhat more common in the fossil record than flowers. Fruits and seeds can be preserved as casts and molds (VG 3:6)(VG 3:7), and fruit o, permineralizations, or less commonly, as three-dimensional organic fossils in clay-rich sediments. Several important seed-rich sites, including the Middle Eocene Clarno Net Beds of Oregon and the Oligocene Brandon Lignite of Vermont have shed substantial light on the ancient angiosperm flora.
|
| Figure 10.14: A collection of Cretaceous flower reconstructions. (A) Chloranthoid flower. (B) Platanoid flower bearing ovules from the Early Cretaceous (Friis & Crepet, 1987). (C) Lesqueria, (D) Archaeanthus and (E) Caloda from the Middle Cretaceous (Dilcher & Kovach, 1986). (F) Prisca (Retallack & Dilcher, 1981), and (G) a generalized reconstruction of the "Rose Creek" flower from the Dakota Groups, Late Cretaceous.
|
![[Previous Page]](../VPLimg/Back.jpeg) |
![[Title Page]](CupD/Cupbutt.jpeg) |
![[Glossary]](../VPLimg/Glossbutt.jpeg) |
![[Range Chart]](../VPLimg/Rangebutt.jpeg) |
![[Geologic Time Scale]](../VPLimg/timesbutt.jpeg) |
![[Next Page]](../VPLimg/Forward.jpeg) |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}