![[Laboratory X - Anthophytes, Glossopterids and Others]](CupD/CupfontB.jpeg)
In 1987, James Doyle and Michael Donoghue published one of the first cladistic analyses including ancient seed plants. This study was designed to look objectively at the relationships among some of the suspected angiosperm "ancestors". From this analysis they recognized the anthophyte clade that included Pentoxylon, bennettitaleans, gnetophytes and angiosperms. Although different analyses have produced somewhat different topologies, most preserve this clade and recognize the gnetophytes as sister to the flowering plants. Synapomorphies for the anthophyte clade include scalariform pitting in secondary xylem, once-pinnate microsporophylls, a single ovule per cupule, derived non-saccate pollen with granular exine structure, syndetocheilic stomata (Figure 10.7), and a tendency to aggregate reproductive structures into flower-like, commonly bisexual, structures.
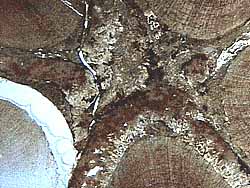
|
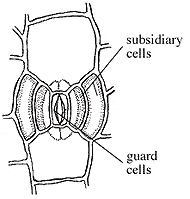
| Figure 10.7: Syndetocheilic stomata in the anthophytes. |
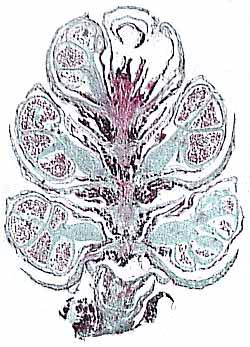
Pentoxylon (Figure 10.8) takes its name from the five wedges of wood that characterize its stem (VG 2:1). Pentoxylon, like Glossopteris and Dicroidium foliages, is a Gondwanan taxon, which has been found in India, Australia and New Zealand. Pentoxylon first appears in the later part of the Paleozoic, but its greatest diversity and abundance appears to be in the Jurassic, continuing into the Early Cretaceous.
|
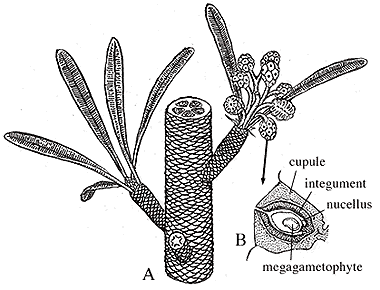
| Figure 10.8: Pentoxylon. (A) Reconstuction of whole plant; note secondary xylem wedges and ovule-bearing structures. (B) Section of oovulate cone in longitudinal section.
|
Pentoxylon has a distinct short-shoot-long-shoot morphology like Ginkgo. Stems are eustelic with secondary wood developing in five or six wedges around a mesarch primary xylem strand (VG 2:1). Note that this eustele arrangement is more like that of Medullosa than that of either conifers or angiosperms. Foliage arises from short shoots, where leaf scars are commonly visible. Leaves a long and narrow with a prominent mid-rib and secondary veins arising at right angles to it and continuing, unbranched to the leaf margin. This foliage type is referred to as Taeniopteris (VG 2:2) and can be distinguished from pinnules of cycads, which can also produce this foliage type, by syndetocheilic stomata (Figure 10.7). In syndetocheilic stomata, the guard cell mother cell gives rise to a pair of guard cells and one or more pairs of subsidiary cells. This contrasts with the haplocheilic condition (Figure 8.6).
Ovule-bearing structures are aggregated into strobili and attached to short shoots (Figure 10.8). Individual cupules contain and almost completely enclose a single ovule. The ovule integument is composed of an inner sclerenchymized layer and a fleshy outer layer. Pollen organs are also born in clusters on short shoots. Note that Pentoxylon does not have bisexual reproductive structures like many other members of the anthophyte clade.
The bennettitaleans (note informal name rather than the order-level Bennettitales) were originally mistaken for cycads because of their cycad-like growth form (Figure 10.9). However, the reproductive structures of these plants and syndetocheilic stomata place them soundly in the anthophyte clade.
|
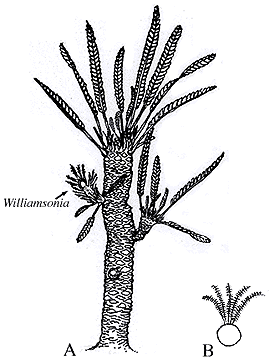
| Figure 10.9: (A) Reconstuction of Williamsonia sewardiana with spirally arranged leaf scars. (B) Cycadeoidea trunk and foliage drawn for comparison of proportions.
|
Bennettitaleans first appear in the Triassic and became extinct toward the end of the Cretaceous. Two major groups are recognized: the cycadeoids with stout trunks and bisporangiate reproductive structures, and relatives of Williamsonia and Williamsoniella, which have slender, branching trunks and either bisporangiate or monosporangiate strobili.
Both groups had woody stems that were heavily armored with persistent leaf bases, much like modern cycads. Both produced entire simple leaves or pinnate foliage (VG 2:3), also like modern cycads. Indeed, the groups differ only in the details of stomatal morphology. This difference, when visible, permits unequivocal segregation of Mesozoic cycad and bennettitalean foliage, such as Nilssonia (VG 2:4). Note that this foliage form genus has also been linked to true cycads. Other foliage-types interpreted as benettitalean include , Anomozamites (VG 2:5) and Zamites (VG 2:6)
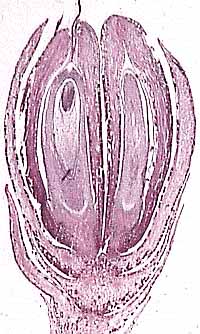
At one point, the bennettitaleans were put forward as possible flowering plant ancestors because of their flower-like bisexual reproductive structures (Figure 10.10). In Williamsoniella, for example, ovules alternate with interseminal scales on an ovulate receptacle at the center of several organ whorls (Figure 10.10a). This reminded some workers of the angiosperm carpel; to others, the complex alternation of interseminal scales and ovule-cupule seemed too derived to be the ancestral state of the angiosperm carpel. The next whorl out was occupied by microsporophylls bearing pollen sacs. This, in turn was surrounded by several whorls of bracts that many homologize with petals and sepals in flowering plants.
|
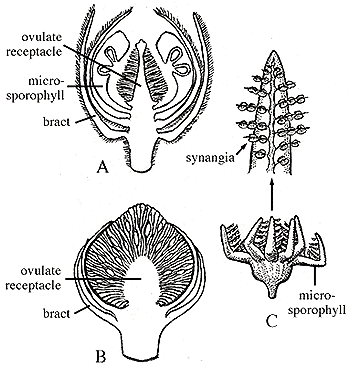
Figure 10.10: Examples of bennettitalean reproductive structures. (A) Williamsoniella bisexual reproductive structure. Note microsporophylls with microsporangia embedded within. (B) Williamsonia ovulate structure. (C) Weltrichia pollen-bearing structure with detail of microsporophyll showing attached synangia. |
Other bennettitaleans bore monosporangiate reproductive structures. Ovulate structures, such as Williamsonia, featured ovulate receptacles, interseminal scales and surrounding bracts, but lacked microsporophylls (Figure 10.10b)(VG 2:7). Pollen organs were composed of a whorl of microsporophylls bearing synangia (Figure 10.10c) (VG 2:8). Note the homoplasy in this character -- bisexual reproductive structures -- that is supposed to be characteristic of the anthophyte clade.
Among living plants, the gnetophytes (Ephedra, Gnetum, and Welwitschia) are believed to be the most closely related to the flowering plants. These living genera also form a monophyletic clade themselves. Like the other extant (non-angiosperm) seed plants, the gnetophytes had their greatest diversity in the past. Today, Gnetum is a tropical moist-habitat plant. In contrast, Ephedra and Welwitschia are both dry-climate or desert plants. Ephedra contains about 30 species with a global distribution, while Welwitschia is represented by a single species that is restricted to some of the driest portions of the Namib Desert in Southwest Africa.
The gnetophytes are characterized by a suite of features that ally them closely with the angiosperms. The leaves of Gnetum, for example, have reticulate venation that bears a striking resemblance to that of some dicot angiosperms. In contrast, Ephedra produces highly reduced spike-like leaves; these are vascularized by a pair of traces that exit from the eustele and enter the leaf. This contrasts with conifers, whose leaves are vascularized by a single trace. Welwitschia has an even more bizarre leaf arrangement in that it produces a single pair of large, parallel-veined leaves that continue to grow throughout the life of the plants. This is a very derived condition that might best be interpreted as progenetic. Also like the flowering plants, the gnetophytes possess vessels in their wood. However, gnetophyte vessels have a subtly different developmental origin and thus may not be strictly homologous with those of angiosperms.
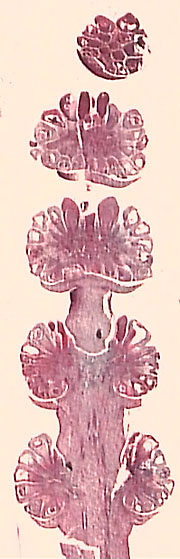
Ephedra is dioecious, with mega- and microsporangiate strobili both being compound structures. Microsporangiate strobili (VG 2:9) consist of several pairs of bracts. A shoot, bearing "bracteoles" (smaller bracts), arises in the axis of each larger bract. Each bracteole surrounds a stalked microsporangiophyll bearing two or more pollen sacs (microsporangia). The megasporangiate cone is similarly arranged with pairs of bracts, the top two of which subtend an ovule (VG 2:10). The ovule is curious in that it appears to be surrounded by two integuments. However, most interpretations call the inner layer the true integument, and the outer layer a reduced bracteole (cupule?) similar to that surrounding the microsporangia.
Ephedra and its sister taxon, Gnetum, are also similar to angiosperms in that they undergoes a form of double fertilization. In most angiosperms, two sperm are involved in fertilization. One unites with the egg cell to form a diploid zygote; the other fuses with the two polar nuclei to form a triploid endosperm. The endosperm serves as nutrition for the developing embryo. In Ephedra, two egg cells are produced, one of which is fertilized to become the embryo while the other begins but does not complete development into an embryo (Friedman, 1994). In Ephedra, this second embryo does not provide nutrition to the developing embryo as does the endosperm of angiosperms; however, the degenerate embryo is believed to be an intermediary step between the normal seed plant condition (where embryo is nourished by a larger gametophyte) and that of angiosperms (megagametophyte so reduced as to not be effective for nourishing the embryo). Another way to think of this is as a reduction in the polyembryony observed in many non-angiosperm seed plants. In some conifers, for example, several embryos form from fertilizations by several pollen grains. Ultimately, only one embryo per seed survives. Clearly, the multi-embryo condition is plesiomorphic for the seed plants and has been reduced in both the gnetophyte and angiosperm lineages.
The reproductive structures of Gnetum are still more flower-like. Strobili are arranged on axes with a conspicuous node-internode organization. Microsporangiate strobili (VG 2:11) have two fused bracts that form a cupule that surround fertile shoots. Each fertile shoot is composed of two fused bracteoles, which surround the microsporophyll. In the megasporangiate strobilus, the cupule subtends a whorl of ovules, which are wrapped in two layers of tissue outside of the integument. These external "envelopes" of tissue may be sclerified and fused to the integument to form a seed-coat-like structure, which makes the Gnetum seed appear very angiosperm-like.
The gnetophytes have a poor fossil record. At present, the earliest reasonable record of the gnetophytes is pollen of Triassic and Cretaceous age, which resembles that of modern Ephedra. The earliest macrofossil gnetophytes appear in the Early Cretaceous, side-by-side with early angiosperm leave of to Potomac Group (Crane and Upchurch, 1987). If we accept Ephedra-like pollen as representative of the gnetophyte clade, then we must conclude that the angiosperm lineage was also independent by the Triassic (see discussion of ghost taxa in Labs 4 and 8). However, we do not have fossil evidence of definitive angiosperms until the Early Cretaceous. Some speculate that early angiosperms lived in habitats so far from likely places of fossil preservation that they elude the fossil record for nearly 100 million years. Another possibility is that although the flowering plant lineage was independent, it had not accumulated the synapomorphies that would allow us to recognize its members as angiosperms.
![[Previous Page]](../VPLimg/Back.jpeg) |
![[Title Page]](CupD/Cupbutt.jpeg) |
![[Glossary]](../VPLimg/Glossbutt.jpeg) |
![[Range Chart]](../VPLimg/Rangebutt.jpeg) |
![[Geologic Time Scale]](../VPLimg/timesbutt.jpeg) |
![[Next Page]](../VPLimg/Forward.jpeg) |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}