|
|
|
|
A case study: Willow Creek Anticline, Two Medicine Formation, Montana |
Science progresses by making observations, asking questions, gathering data, and testing hypotheses. However, these steps do not necessarily proceed in a linear fashion. Along the path of scientific exploration, scientists re-test hypotheses, make new discoveries, add new data, ask new questions, and use new technology that allows for innovative ways of testing hypotheses. Over time, additional data and research either strengthen or weaken original hypotheses. The research on eggs and eggshell from the Two Medicine Formation is an example of the often circuitous scientific process.
| Click any image on this page to see an enlargement. | |
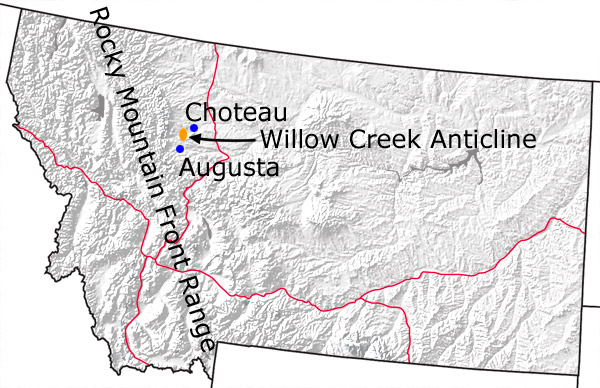 Figure 1. Map of Montana with the area containing the Willow Creek Anticline (including Egg Mountain and Egg Island) in orange.  Figure 2. Typical badlands outcrop of the Two Medicine Formation in the Willow Creek Anticline. |
Exploration and discovery
Initial observations
Egg Mountain and the nearby nesting sites in the Two Medicine Formation have
produced fossil remains of embryonic, hatchling, juvenile, and adult dinosaur skeletons, as well as eggs, eggshell fragments, and nest structures. Several
dinosaur species have been found, including Maiasaura peeblesorum, Orodromeus
makelai, and Troodon formosus. Skeletons of lizards, turtles, and
pterosaurs have also been found. At least four eggshell morphotypes are
present, either as isolated fragments, within nest structures, and/or associated
with embryological or hatchling skeletal material. The sediments in which the
eggs and dinosaur fossils are found represent stream and lake deposits that
were adjacent to the young, uplifting Rocky Mountains. Ash deposits are present
in the area, indicating nearby volcanism.
Asking questions
The initial observations of eggs, nests, baby dinosaurs, and adult dinosaurs have led researchers to ask many questions regarding the biology and behavior of dinosaurs discovered at nesting sites within the Willow Creek Anticline. Some of the questions they have asked include:
Which eggshell types are present and which are associated with embryonic remains in the Willow Creek Anticline? How do these eggs compare to eggs from other areas (both within the Two Medicine Formation and throughout the world)?
What does the association of eggs, nests, and baby dinosaurs tell us about dinosaur reproductive behavior?
Given the presence of embryonic, hatchling, juvenile, and adult skeletal material at Egg Mountain, what can researchers learn about the ontogeny (growth through time) of various dinosaur species?
What does the position of eggs within nests indicate about the reproductive biology and behavior of these dinosaurs?
Can eggshell, nest structures, and dinosaurian growth rates tell us anything about the evolutionary transition from dinosaurs to birds?
Gathering and interpreting data
Studies on the fossils collected from the Willow Creek Anticline nesting sites
demonstrate the breadth of information that can be gained from the discovery
of skeletal fossils in association with nests and eggs. Over the three decades
since the original discoveries were made, scientists have put together a story
of the dinosaurs that lived in the Late Cretaceous Two Medicine Formation that
is continually being refined and updated.
Maiasaura nests and eggs
After the first discoveries of egg and skeletal fossils, scientists' initial
hypotheses focused on dinosaur nesting and parental behavior. The first publication
on Two Medicine Formation nesting sites described eggshell fragments and nestling Maiasaura
peeblesorum bones that were found within and around a nest-like structure.
The partially-grown nestlings and associated nests and eggshell were interpreted
as evidence that the babies remained in the nest after hatching, and led Horner
and Makela (1979) to hypothesize that this was evidence for parental care.
Horner proceeded to uncover and study more egg and nest localities in the area.
Not only did he find multiple nests on the same depositional horizon — evidence
suggesting colonial nesting behavior — but he also found multiple horizons
of nests layered one on top of each other. From this, Horner concluded that groups
of dinosaurs returned to the same area over multiple breeding seasons (Horner 1982).
Troodon nests and eggs
After the initial descriptions and behavioral interpretations of Maiasaura nesting
activity, researchers began to study the different types of eggs and eggshells
collected from nearby Egg Mountain and Egg Island. These eggs were found at
different stratigraphic levels, were more elongate with a different surface
pattern than the Maiasaura eggs, had a different microstructure,
and were found with distinctly different embryonic remains. The eggs and embryos
were eventually identified as the small theropod dinosaur Troodon. With
two species of dinosaur directly associated with eggs and eggshell fragments,
growth, behavior, and reproductive physiology could be compared between different
species.
Ontogeny
Ontogeny is the study of how organisms change over time — that is, the
study of growth. Ontogenetic investigations are most informative when an entire
growth series is available for analysis — embryos, hatchlings, juveniles,
and adults. The discovery of embryonic, hatchling, and adult remains at the
Willow Creek Anticline localities provided material for ontogenetic comparisons.
Horner and his colleagues compared the skeletal characteristics of Maiasaura and Troodon embryonic
remains to determine the likely post-hatching behavior and locomotion (Horner
and Weishampel 1988, 1996). Differences in bone development (ossification)
between the two dinosaur species led the researchers to draw a connection between
growth rate and assumed parental behavior for both species of dinosaurs. Comparisons
of the leg bones revealed that those from Maiasaura were not fully ossified,
whereas, those from Troodon were. These findings led Horner and Weishampel
to hypothesize that Maiasaura received prolonged parental care in the
nest, and that Troodon hatchlings were able to leave the nest immediately.
In addition, later histological studies of embryonic and hatchling bones showed
that Maiasaura grew more slowly than Troodon (Horner et al. 2001),
supporting the original hypotheses.
Reproductive physiology
 Figure 2. A cast of Troodon formosus eggs and nest structure from the Egg Mountain locality in the Two Medicine Formation of central Montana. UCM 239, photograph CO96.18.  Figure 3. Three of the eggshell morphologies discovered in the Willow Creek Anticline. Two of the eggshells, (A) and (B), were found with associated embryonic material, so the eggshell can be assigned to the species of dinosaur that laid the egg. A. Dinosauroid Prismatic eggshell of Troodon formosus. Specimen UCM 239. B. Dinosauroid Spherulitic (specifically Dinosauroid Prolatospherulitic) eggshell of Maiasaura peeblesorum. Specimen UCM 365-2, photomicrograph M729.4A (x50). C. Ornithoid Ratite eggshell. Specimen UCM 240-R. |
When whole eggs are preserved within nest structures, scientists have an opportunity to study how organisms built nests and laid eggs. Studying the relationship of Troodon nests and eggs (Fig. 3), Varricchio et al. (1997) identified aspects of reproductive physiology and behavior based on the orientation of the eggs. Statistical analysis indicated that eggs were paired in the nest (two eggs were closer to each other than to neighboring eggs), implying that eggs were laid two at a time. Only an organism with two functioning oviducts can do this. However, the asymmetric shape indicates Troodon only retained one egg in each oviduct at a time, presenting a combination of characteristics not found in extant amniotes: two functioning oviducts with one egg in each oviduct at a time. Because the eggs were only partially buried by sediment in the nest, the authors also concluded that Troodon likely brooded their eggs, providing additional warmth for the eggs and developing embryos. These features indicate that both bird-like and crocodilian-like characteristics were present in Troodon breeding behavior and reproductive anatomy. Primitive (or non-birdlike) features include two functioning oviducts, lack of egg rotation, and partial burial of eggs with sediment (which precludes egg rotation). Derived (or birdlike) features include large eggs, one egg formed in each oviduct at a time, and brooding behavior (hypothesized to have been used in conjunction with the warmth of the sediment for egg incubation). The unique opportunity to study dinosaur reproduction adds important information to the analysis of the evolutionary relationships between non-avian theropods like Troodon and birds.
Eggshell morphology
Because embryonic remains were preserved with eggshell, scientists were able
to correlate eggshell morphology with
the genus or species of animal that laid the egg. Hirsch and Quinn (1990) studied
eggshell morphology and microstructure from Egg Mountain and surrounding areas
and described four morphotypes — dinosauroid prismatic, dinosauroid spherulitic,
and two ornithoid morphologies (Fig. 4; see "Eggshell
differentiation and identification" for
more information on morphotypes). This information has given scientists important
insights on similar fossil eggshell material that was not associated with embryonic
remains.
Cladistic analysis
Although the evolutionary relationship between non-avian theropods and birds
has been a hot topic, an overwhelming majority of scientific evidence now supports
the hypothesis that birds
evolved from bird-like theropod dinosaurs. The preservation of eggs and associated
embryonic material found at sites around the Willow Creek Anticline has added
data in support of this evolutionary relationship. Varricchio and Jackson (2004)
used eggshell morphologies of turtles, crocodilians, dinosaurs, and birds to
perform phylogenetic analyses looking for patterns in eggshell characters among taxa. Troodon eggshell
and an unnamed eggshell morphology from the Two Medicine Formation were found
to be very similar to bird eggshells. These results support the close affinity
of theropod dinosaurs and modern birds and bolster arguments for the theropod
dinosaur origin of birds. The study also provided evidence for shared reproductive
characteristics between Troodon and avian theropods (i.e., birds). The cladistic approach
using discrete external and internal structural characters provides a new direction
for eggshell research and breaks away from the historic approach to eggshell
classification based on morphotypes.
Testing ideas and community analysis
After initial hypotheses are proposed, these hypotheses are often tested and
re-tested with additional data and observations. When new data or analytical
techniques can supplement original studies, the initial hypotheses can be revised,
or different interpretations of data can lead to new, opposing hypotheses. Research
on the Willow Creek Anticline nesting sites has followed this process. Over the
years, hypotheses have been challenged, revised, or supported with additional
scientific research and data collection. It should be noted that opposing hypotheses
are not necessarily more accurate simply because they are newer; they often just
provide a different way of interpreting the data.
Revised hypothesis
Perhaps one of the most significant revisions to the original Egg Mountain hypotheses
came from two of the initial researchers. In 1996, Horner and Weishampel corrected
their original conclusion (Horner and Weishampel 1988) that the elongate, dinosauroid
prismatic eggs were laid by the small hypsilophodontid Orodromeus makelai.
Further research and preparation of the eggs and embryological material provided
more data allowing for reanalysis — the eggs, it turned out, contained Troodon
formosus embryos. This revised conclusion has had a considerable effect on
the original interpretation of dinosaur nesting behavior, community structure,
and eggshell morphologies. Since the eggs and eggshell structures are similar
to avian eggs, the fact that the embryological remains and associated eggs were
attributed to a theropod has major implications for interpreting non-avian theropod
reproductive physiology.
Opposed hypotheses
In their original description of embryonic remains from the Willow Creek Anticline,
Horner and Weishampel (1988) cited degree of ossification of the leg bones
of Maiasaura and Troodon (then thought to be Orodromeus)
to indicate the level of mobility of young after hatching. Subsequently, Geist
and Jones (1996) compared extant perinatal (the developmental stage
immediately prior to and following hatching) birds and crocodilians to fossil
dinosaur embryos and hatchlings. They found that the extent of hip bone development
was more important than leg bone development for recognizing precocial versus altricial hatchlings,
and that the leg bones of Maiasaura, Troodon, and other dinosaurs
did not reliably indicate the mobility of a hatchling. Geist and Jones suggested
that the hatchling dinosaurs studied were likely precocial upon birth, although
this does not preclude the provision of extended parental care. Horner et al.
(2001) countered Geist and Jones' (1996) argument after an extensive
histological analysis of turtle, crocodilian, non-avian dinosaur, and bird
embryonic and perinatal bones that compared bone developmental patterns and
growth rates. The authors correlated ossification and growth rates with
life-history strategies. Horner et al. (2001) concluded that developmental
differences (including growth rates) in embryonic and perinatal dinosaur bones
from the Willow Creek Anticline indicate a precocial lifestyle for Troodon and Orodromeus hatchlings
and an altricial lifestyle for hadrosaur hatchlings that necessitated parental
care; this work supported their original hypothesis (Horner and Weishampel
1988).
The original hypothesis regarding the spatial arrangement of eggs in nests and the physiological and behavioral implications of this arrangement has also been reviewed. After the initial clutch analysis by Varricchio et al. (1997), other researchers (Grellet-Tinner et al. 2006) questioned the laying of Troodon eggs in pairs, citing possible post-depositional alteration of the spatial arrangement of eggs and statistical questions. Grellet-Tinner et al. (2006) proposed that Troodon laid unpaired eggs and thus had one functioning ovary, rather than two as concluded by Varricchio et al. (1997). However, this second hypothesis lacks the direct fossil evidence of the two-functioning-ovary hypothesis (Varricchio et al. 1997).
Outcomes and feedback
The effects of these discoveries on the scientific community and general public
are difficult to gauge, but unique discoveries undoubtedly increase enthusiasm
and rejuvenate interest in various aspects of dinosaur paleontology. The Two
Medicine Formation nesting site discoveries and subsequent research have added
to the already increasing energy surrounding debates over dinosaur physiology
(e.g., the warm-blooded/cold-blooded debate), evolutionary relationships between
dinosaurs and birds, and dinosaur behavior. The focus on eggs, eggshell fragments,
and associated embryological material has also helped to rejuvenate scientific
interest in fossil eggshell and the paleobiological,
paleoenvironmental, and behavioral information contributed by eggshell research.
Few nesting sites with embryonic and juvenile skeletal material were known
prior to 1978, so the numerous studies on the fossil eggs, eggshell, and juvenile
dinosaurs from the Willow Creek Anticline have provided a foundation on which
new scientific ideas are being built.
Additional readings
Geist, N.R., and T.D. Jones. 1996. Juvenile skeletal structure and the reproductive habits of dinosaurs. Science 272: 712-714.
Grellet-Tinner, G., L. Chiappe, M. Norell, and D. Bottjer. 2006. Dinosaur eggs and nesting behaviors: A paleobiological investigation. Palaeogeography, Palaeoclimatology, Palaeoecology 232:294-321.
Hirsch, K.F., and B. Quinn. 1990. Eggs and eggshell fragments from the Upper Cretaceous Two Medicine Formation of Montana. Journal of Vertebrate Paleontology 10(4):491-511.
Horner, J.R. 1982. Evidence of colonial nesting and 'site fidelity' among ornithischian dinosaurs. Nature 297(5868):675-676.
Horner, J.R. 1984. The nesting behavior of dinosaurs. Scientific American 250(4):130-137.
Horner, J.R., K. Padian, and A. de Ricqlès. 2001. Comparative osteohistology of some embryonic and perinatal archosaurs: Developmental and behavioral implications for dinosaurs. Paleobiology 27:39-58.
Horner, J.R., and R. Makela. 1979. Nest of juveniles provides evidence of family structure among dinosaurs. Nature 282(5736):296-298.
Horner, J.R., and J. Gorman. 1988. Digging Dinosaurs. Workman Publishing, New York. 210 pp.
Horner, J.R., and D.B. Weishampel. 1988. A comparative embryological study of two ornithischian dinosaurs. Nature 332:256-257.
Horner, J.R., and D.B. Weishampel. 1996. A comparative embryological study of two ornithischian dinosaurs — A correction. Nature 383:103.
Varricchio, D.J., F. Jackson, J.J. Borkowski, and J.R. Horner. 1997. Nest and egg clutches of the dinosaur Troodon formosus and the evolution of avian reproductive traits. Nature 385:247-250.
Varricchio, D.J., F. Jackson, and C.N. Trueman. 1999. A nesting trace with eggs from the Cretaceous theropod dinosaur Troodon formosus. Journal of Vertebrate Paleontology 19(1):19-100.
Varricchio, D.J., and F.D. Jackson. 2004. A phylogenetic assessment of prismatic dinosaur eggs from the Cretaceous Two Medicine Formation of Montana. Journal of Vertebrate Paleontology 24(4):931-937.
 |
|
 |
|
 |
|
 |
| Fossil eggshell home | Karl Hirsch and the Hirsch Eggshell Collection | Case studies | Interactive map |
Shaded relief map from the USGS, adapted by The Paleontology Portal (www.paleoportal.org); Figure 2 © Karen Chin; Figures 3 and 4 courtesy of the University of Colorado Museum.