Tracking the Course of EvolutionEVOLUTION AND SYSTEMATICS (cont.)by Sandra J. Carlson
METHODS OF PHYLOGENY RECONSTRUCTION
1. Choosing the Taxa
2. Selecting Characters
3. Deciding a Point of Reference
4. Selecting the Method
5. Phylogenetic Analysis
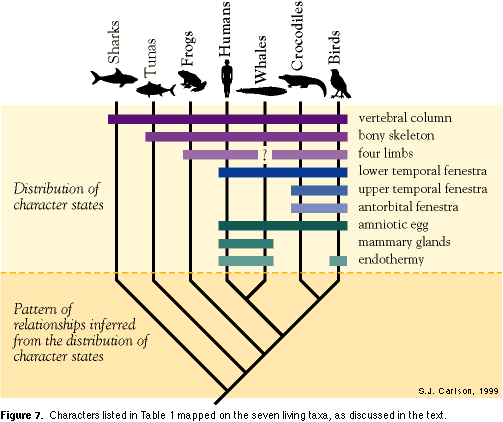
Character 1. Vertebral column. As their name reveals, vertebrates all have a vertebral column as the central component of their skeleton. All members of the ingroup are vertebrates, and the two outgroup taxa chosen are also from the vertebrate clade. The fact that all the ingroup and outgroup taxa share a vertebral column indicates that they share more recent common ancestry with each other than any one taxon does with a non-vertebrate animal. Thus, sharks and tuna are more appropriate outgroup taxa for a group that includes frogs, humans, whales, crocodiles, and birds than are, for example, clams or insects, which both lack a vertebral column and are more distantly related to vertebrates. Character 2. Bony internal skeleton. Tuna share with the ingroup taxa an internal mineralized skeleton composed of calcium phosphate (bone), while sharks have an internal skeleton composed entirely of non-mineralized cartilage, not bone. Because bone develops in the same way, has the same chemical composition, and is located in the same region of the body in all bony vertebrates, bone is considered to be a homologous character. The distribution of bone among the ingroup and outgroup taxa indicates that the five ingroup taxa share more recent common ancestry with the tuna than with the shark. Character 3. Four limbs. Frogs, humans, crocodiles, and birds have two forelimbs and two hindlimbs; they are all tetrapods. Birds have wings instead of "arms" as forelimbs, but they are still tetrapods. Whales are somewhat confusing, because they have become fully aquatic secondarily, a habitat that their fully terrestrial ancestors successfully invaded and adapted to millions of years ago. Their forelimbs have been modified to flippers, and hindlimbs have been lost entirely (at least in living whales). Since we can't observe two hindlimbs in living whales, I have indicated with a question mark the state of this character in Table 1. Before doing an analysis, we can't be sure if they lost their hindlimbs secondarily, or never had hindlimbs at all. Sharks and tuna both have fins that are fundamentally different from limbs, even those present in whales, revealing fins as the ancestral state of this character. Having four limbs is the derived state of Character 3, and the four tetrapod taxa that share the derived state are thus united in a clade.
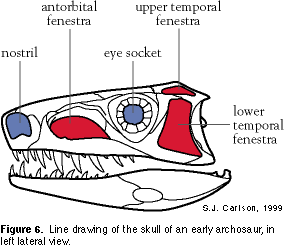
Character 4. Lower temporal fenestra. The number and position of openings in the skull (fenestra) within and between bones are diagnostic characters, particularly among the tetrapods. From a side view of the skull, mammals (humans and whales) possess a lower temporal fenestra — a single opening below and behind the eye socket (Figure 6). Even extinct relatives of mammals, including Dimetrodon, possess this skull opening. Crocodiles and birds possess an upper temporal fenestra, in addition to a lower temporal fenestra (see below). Sharks, tuna, and frogs have skulls, but do not possess openings in the same positions on the skull that other tetrapods do; they lack a lower temporal fenestra, which is the ancestral state of this character.
Character 7. Amniotic egg. Of our five ingroup taxa, only humans, whales, crocodiles and birds have a protective membrane, called an amnion, surrounding their eggs. The amnion allows the egg to survive outside of an aquatic environment. In both humans and whales, the egg develops inside the mother's body, but still possesses an amnion. Frog (and shark and tuna) eggs are different from human, whale, crocodile, and bird eggs; they lack this membrane, which is the ancestral state of the character, and must lay their eggs in water if they are to survive.
Character 8. Mammary glands. All living mammals, including humans and whales, possess mammary glands to nurse their young. No other living vertebrates share this derived character, which is clearly related to the mammalian mode of reproduction.
RETURN TO TOP
Character 9. Endothermy. Endothermy is the ability to regulate body temperature internally, rather than rely on the external environment to warm up or cool down body temperature passively (a condition referred to as ectothermy). Humans, whales, and birds are all endothermic and their body temperature doesn't vary by more than a degree or two throughout the day. Sharks, tuna, and frogs are all ectothermic. This is the ancestral state of this character, as determined by its pattern of distribution relative to Characters 1 through 4. Because crocodiles also share the ectothermic condition, we infer that they retain it from their ancestors. Therefore, we infer that endothermy evolved two times independently, once in mammals (humans and whales) and once in birds. In this analysis, endothermy is a homoplastic character, and by itself might falsely lead us to conclude that mammals and birds are more closely related to each other than either is to crocodiles. Based on the distribution of a larger suite of characters that are in conflict with this interpretation (including Characters 5 and 6), we can detect endothermy as a homoplastic rather than homologous character, at this level of analysis. Characters 10 and 11. Reduced fourth and fifth digits on hands; fully upright posture. A shared derived character uniting tetrapods (in addition to two arms and two legs), is the presence of five fingers and five toes. In frogs, humans, and crocodiles, all five fingers are of more or less similar length. In birds and both saurischian and ornithischian dinosaurs, the fourth and fifth digits (ring finger and pinky finger) are reduced considerably relative to the other fingers. Birds and both groups of dinosaurs also share a fully upright posture, with the hindlimbs approximately parallel to the vertebral column. Even though they are extinct, we can infer posture in terrestrial dinosaurs from footprints and from the shapes of the joints between limb bones, particularly the hip, knee, and ankle joints. Crocodiles have a semi-erect posture, and frogs have a squat, sprawling posture, with their limbs emerging at an angle from the vertebral column. Humans and other mammals (with hind limbs, so not including whales) have evolved an upright posture independently from that in birds and dinosaurs, similar to the evolution of endothermy in both mammals and birds.
Characters 12 and 13. Long S-shaped neck; long hands. Birds and the saurischian dinosaurs both have relatively long legs, arms, and hands, and long S-shaped necks, while crocodiles have much shorter legs, arms, hands, and necks relative to body length. This is considered to be the ancestral condition, since crocodiles lie outside the clade containing birds and dinosaurs, based on the distribution of Characters 10 and 11 (and many others).
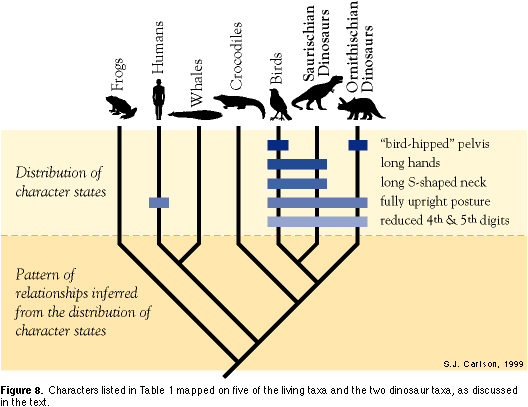
As is true for each of the nodes in these cladograms (Figures 4, 7, 8), numerous other characters can be listed to provide support, but are not discussed for lack of space. However, since the saurischian and ornithischian dinosaurs are known most commonly as the "lizard-hipped" and "bird-hipped" dinosaurs, it is worth mentioning the distribution of pelvic structure on this cladogram (Figure 8), because it appears to be in conflict. Character 14. "Bird-hipped" pelvis. Saurischian dinosaurs share the ancestral "lizard hipped" condition with crocodiles (and, of course, lizards). Birds and ornithischian dinosaurs have each evolved a "bird-hipped" pelvic structure independently from this ancestral "lizard-hipped" condition, a conclusion that is supported by many other additional skeletal features. The character is thus homoplastic, in this example. Taking all the information on character distribution in the ingroup and comparing it to the states present in the outgroup, we can construct the cladograms illustrated in Figures 7 and 8. The shared derived characters that support each node (and, consequently, each clade nested within each more inclusive clade) are listed beside them. On the basis of this phylogenetic hypothesis, we can make the evolutionary statement that birds and other dinosaurs share more recent common ancestry than either group does with crocodiles, or with mammals, or sharks. This is a powerful evolutionary statement, and a very useful one, if we agree that our goal in systematics is to discover the pattern of evolution. Simply stating that birds are in the Class Aves, dinosaurs in the Orders Ornithischia and Saurischia, and crocodiles in the Order Crocodylia emphasizes each group as a unit separate from the others, rather than as elements in a clade that are related by common ancestry. |
| Return to top of page | TRACKING HOME | PRESENTATIONS |