Tracking the Course of Evolution
EVOLUTION AND SYSTEMATICS (cont.)
NOTE: This is page 4 of a five-page document.
SYSTEMATICS AND THE FOSSIL RECORD
Life on Earth in the past was different in many ways from life on Earth today. Every new fossil discovered further supports this claim. Working through the dinosaur example above, I hope I have convinced you that systematics, as a method for inferring evolutionary relationships, can be applied to both living and fossil organisms. Does fossil data require any special consideration in systematic investigations? The evidence of evolution from the fossil record particularly relevant to systematics comes in three forms: shape, space, and time.
Shape
Perhaps the most valuable contribution that the fossil record provides is a tangible record of extinct fossil morphologies or character combinations that no longer coexist in organisms today. These unique character combinations are invaluable in piecing together relationships among living organisms that look quite different from one another (birds and crocodiles, for example). However, fossil data comes almost exclusively in the form of morphology, usually skeletal morphology, which we must assume bears some interpretable relationship to the underlying genome, since the genome in fossils cannot be sampled directly. Aspects of morphology that are not mineralized are less likely to be preserved as fossils, although some soft tissues (muscles, for example) can leave an interpretable scar or mark on skeletal fossil material. Developmental or physiological processes of extinct organisms cannot be observed directly or experimented with. Although we can infer some types of behavior from trace fossils, we can't observe fossils locomoting, eating, reproducing — in other words, living. In this sense, fossils provide us with a much-reduced suite of characters for systematic analysis than do living organisms.
Space
Because of plate tectonics, the geography of the world today is different than it was in the past, and has changed continually over geologic time. Thus, we might find fossil organisms from different continents, or from sediments indicating different paleoclimatic conditions than where closely related living organisms are found today. For example, close relatives of camels and elephants are found as fossils in North America, but unfortunately we don't have camels and elephants living in North America today. Fossils can provide the only tangible record of ancient biogeographic ranges and can fill in important paleogeographic gaps between disjunct modern occurrences of closely related species. Paleobiogeographic data from fossils can thus be used to test hypotheses of phylogenetic relationship, and patterns of dispersal, among living taxa.
Time
The fossil record provides an ordered temporal record of the relative appearance of unique character combinations over millions of years of geologic time. What the fossil record lacks in direct evidence of development, genetics, physiology, behavior, and ecology, it provides in abundant sequential data on skeletal morphology that cannot be obtained from any other source. Characters hypothesized to exist in a common ancestor on the basis of their distribution among living taxa can be tested by their discovery at any of a number of stratigraphic levels, ordered in time. A relative temporal pattern implied by the internesting of clades one within another (Figure 4) can be tested by its correspondence to the relative appearance of fossils, and the characters they possess, in a stratigraphic sequence (Fisher, 1991; Huelsenbeck, 1994; Benton, 1995; Wagner, 1995). Hypotheses of character homology may also be tested (for example, the evolution of the bones of the inner ear in amniotes). Some argue that character polarity can be read directly from the relative appearance of characters in the fossil record; character states that appear earlier are primitive, those that appear later are derived. Arguments about the incompleteness of the fossil record and nature of sampling from the fossil record are relevant to this issue, but are discussed in more detail in other chapters of this book and will not be revisited here.
RETURN TO TOP
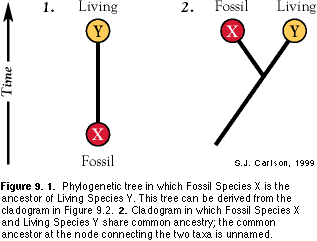
 The fossil record was once viewed (and is still by some scientists) as the archive of ancestors. Examining the distribution of character states among living organisms in Species Y and extinct organisms in Fossil Species X, one could hypothesize a direct ancestor-descendant evolutionary relationship, diagrammed as an evolutionary tree (Figure 9.1). The assumption that all (or most) ancestors of living taxa are preserved as actual fossils (see Foote, 1996) is a difficult assumption for many phylogenetic systematists to accept. Why is this so? In addition to the completeness issues raised above, the following argument is relevant. If Fossil Species X is distinct enough in morphology to be given its own species name (which is most often the case), then this indicates that some evolutionary change has occurred between X and its true ancestor, which is really the common ancestor of both X and Living Species Y (Figure 9.2). Here, X is hypothesized to share common ancestry with Y, but is not necessarily considered to be the true ancestor of Y for the sole reason that it occurs earlier in time. The pattern of common ancestry diagrammed in Figure 9.2 is a more conservative statement of relationship, requiring fewer assumptions, than one of direct ancestry, and is preferred for that reason. The fossil record can be viewed as a source of morphological characters that can better reveal patterns of common ancestry, and not only as a source of true ancestors. The fossil record was once viewed (and is still by some scientists) as the archive of ancestors. Examining the distribution of character states among living organisms in Species Y and extinct organisms in Fossil Species X, one could hypothesize a direct ancestor-descendant evolutionary relationship, diagrammed as an evolutionary tree (Figure 9.1). The assumption that all (or most) ancestors of living taxa are preserved as actual fossils (see Foote, 1996) is a difficult assumption for many phylogenetic systematists to accept. Why is this so? In addition to the completeness issues raised above, the following argument is relevant. If Fossil Species X is distinct enough in morphology to be given its own species name (which is most often the case), then this indicates that some evolutionary change has occurred between X and its true ancestor, which is really the common ancestor of both X and Living Species Y (Figure 9.2). Here, X is hypothesized to share common ancestry with Y, but is not necessarily considered to be the true ancestor of Y for the sole reason that it occurs earlier in time. The pattern of common ancestry diagrammed in Figure 9.2 is a more conservative statement of relationship, requiring fewer assumptions, than one of direct ancestry, and is preferred for that reason. The fossil record can be viewed as a source of morphological characters that can better reveal patterns of common ancestry, and not only as a source of true ancestors.
 CLASSIFICATION
CLASSIFICATION
No discussion of evolution and systematics would be complete without considering classification. Classification refers to the ways that we organize organisms. Human beings name organisms (and most other objects) in the world. Names identify individuals uniquely; for example, Lou Gehrig was a particular baseball player. In the binomial system a name refers to both the particular individual and his or her family ancestry; Lou was a member of the Gehrig family. In some cultures, each person has a single unique name, like Confucius or Sacajawea. In Mexico, individuals might retain both their mother's and father's names; Maria Fernandez Chacon could be the daughter of José Fernandez and Juanita Chacon. In China, the surname is first; Sun is the family name of Sun Yat-Sen.
Just as we refer to each other by a given name and a surname, the international scientific community refers to organisms by a species name and a genus name, listing the genus name first. Consider the familiar dinosaur, Triceratops horridus. Triceratops is the genus name that means "having three horns"; horridus is the species name that means "projecting" or "standing on end" (rather than horrid, as you might expect!) and refers to the orientation of the horns on the skull. By convention, genus and species names are always italicized or underlined. Scientific names often, but not always, refer to some feature or special characteristic of the organism that may helps us remember what a given species looks like, and how it differs from other species.
RETURN TO TOP
Too many species exist today and throughout the fossil record for us to remember and communicate about each one individually. We need to agree on a set of rules for naming groups of species (and groups of groups of species), to make it easier for us to talk about them. Classification refers to a system of organizing and naming living (or once living) things. What rules should we use for classifying organisms? By now, the usefulness of identifying phylogenetic relationships among species and clades should be clear — evolutionary processes structure the diversity of life and phylogenetic methods may be used to reconstruct the pattern of evolution. For this reason, it is most informative to name groups of species on the basis of their phylogeny. Recall that phylogenies are hierarchical and clades are internested (Figure 4). Naming successively more inclusive clades, or groups of species that share common ancestry, reflects the process of descent with modification.
Not all classifications are organized according to patterns of phylogenetic relationships, however. In fact, the system of classification that biologists and paleontologists use today was established before evolution was recognized as the process structuring patterns of phylogeny. (See Hedegaard, this volume.) In the eighteenth century, a Swedish biologist named Carl Linné developed a system for naming groups of species (higher taxa) and ranking them in a particular hierarchy or taxonomy (Figure 10) quite separate from evolutionary theory.
 Because evolutionary principles do not structure the Linnean system of classification, some, but not all named higher taxa are true clades. This can present a problem when higher taxa are treated uncritically as if they were clades. Phylogenetic systematists are working to identify the phylogenetic status of taxa named under the Linnean system. The grouping of genera into families, families into orders, and so on, are being revised to better reflect patterns of evolution. Some systematists have proposed a complete overhaul of the classification system itself so the principles governing the grouping of species into higher taxa are based fundamentally on evolution (de Queiroz and Gauthier 1990, 1992). Perhaps by the next millenium, the ways in which systematists classify organisms will be based on truly evolutionary criteria.
Because evolutionary principles do not structure the Linnean system of classification, some, but not all named higher taxa are true clades. This can present a problem when higher taxa are treated uncritically as if they were clades. Phylogenetic systematists are working to identify the phylogenetic status of taxa named under the Linnean system. The grouping of genera into families, families into orders, and so on, are being revised to better reflect patterns of evolution. Some systematists have proposed a complete overhaul of the classification system itself so the principles governing the grouping of species into higher taxa are based fundamentally on evolution (de Queiroz and Gauthier 1990, 1992). Perhaps by the next millenium, the ways in which systematists classify organisms will be based on truly evolutionary criteria.
BACK to page 3
CONTINUE to page 5
|