![[Lab VI - Sphenopsids and Ferns]](SpheFeD/SpFefontA.jpeg)
Until some of these systematic issues discussed above are better resolved, we will give a few examples as an overview of early fern representatives.
Rhacophyton. Rhacophyton is well known from the Late Devonian of North America, Europe and western Siberia. It was characterized by a semi-erect axis with rows of paired branches. Branches may have two lateral rows of alternate pinnae that form a branch system in a single plane (biseriate) or they may produce a more three-dimensional arrangement of pinnae. Although the biseriate branches look much like the fronds of modern ferns they are not. Fronds are, strictly speaking, elaborate compound leaves, attached to the stem by a petiole. The frond-like structures of Rhacophyton are clearly branches with secondary xylem. Homosporous sporangia of Rhacophyton are borne on three-dimensionally branched appendages; sporangia are fusiform and terminal.
The discovery of secondary xylem in the frond-like structures of Rhacophyton illustrate the taxonomic dilemma posed by this plant. If you saw it growing today, you probably wouldn't recognize it as a fern. However, its stele morphology is like many other things that we recognize as ferns. The branching rather than frond architecture led Andrews and Phillips (1968) to conclude that it was more closely allied with the progymnosperms. Cornet and colleagues (1976) argued that Rhacophyton, while neither fern nor progymnosperm, could "serve as a focal point for fern-like plants which may have progymnosperm characters but are apparently not in a direct line of gymnosperm evolution." This taxon, therefore, illustrates two important points about the origin and early evolution of ferns: (1) the origin of this important group remains obscure; and (2) the lack of clear synapomorphies that unite all the things that we call ferns (even the modern ones) may suggest evolution of a variety of plants out of an ancient complex of plants that populate the base of this radiation. Confused? Read on!
Stauropteris. The genus Stauropteris is known from the Carboniferous (Mississippian and Pennsylvanian) of Europe and North America. Elaborate three-dimensional branching systems are well on their way to becoming fronds, with their vascular strands distinctly different from that of the rhizome. The rachis is characterized by a four-lobed mesarch protostele (VG 2:7). Primary xylem strands dichotomize to generate the tetrarch vascular strand associated with successive ranks of pinnae. At a higher level on the rachis and on the opposite side, a second pair of traces is produced. Stauropteris was heterosporous and produced eusporangiate megasporangia terminally on ultimate branches. Megasporangia contained two functional megaspores; microsporangia appear to have been borne on different individuals (often identified as different form species).
A phylogenetic analysis by Rothwell (1996) places this genus sister to the Early Devonian plants Psilophyton and Aglaophyton. This topology means that these are not closely related to ferns at all, even though much of the previous literature and evolutionary thinking has placed them among the possible fern ancestors. You'll note that we've conveniently left Stauropteris (and the genera discussed in the next paragraph) out of the overall phylogeny in the Virtual Paleobotany Laboratory. This choice respects continued debate about relationships of these groups to other lineages of vascular plants. However, we discuss Stauropteris (and Zygopteris and friends, below) in this laboratory because of their more traditional interpretation as possible fern relatives.
Zygopteris, Etapteris, Metaclepsydropsis, and Clepsydropsis. The genera Zygopteris (Carboniferous-Permian), Etapteris (Carboniferous), Metaclepsydropsis (Lower Carboniferous), and Clepsydropsis (Lower Carboniferous), traditionally united in the "Zygopterids", were important components of the flora from the Late Devonian into the Permian They all have quadriseriate fronds in which pinnae are borne at right angles to each other rather than in a single plane (known as biseriate) as in most modern ferns. The main axis is a protostele, with rachis traces arising in a spiral. The rachis trace is often complexly lobed and may be "H-shaped" or "hour-glass-shaped" (VG 2:8). The pinnae traces arise from the rachis trace in pairs that depart from the ends of the H-arms. Ultimate pinnae have dichotomizing veins. Fertile fronds have clusters of elongate sporangia that partially replace pinnules. Sporangia may have one or several wall layers, making them appear transitional between eusporangiate and leptosporangiate forms. In fact, their mix of characters beyond the sporangia complicate this gradualist interpretation.
Rothwell's (1996) phylogenetic analysis returns these genera (along with Rhacophyton) sister to the Progymnosperms and seed plants. Since this topology depends on a number of key homologies that are still up for debate, we have not included these plants in the overall phylogeny presented in the Virtual Paleobotany Laboratory. This hypothesis is interesting and bears careful consideration. However, such an interpretation would unseat these plants as possible ancestral fern morphologies. Such an interpretation would leave fern origins even more obscure.
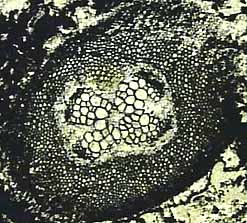
Marattiales. The Marattiales seems to be a good phylogenetic group that includes some living and a variety of ancient forms that share many synapomorphies. These are uncommon tree ferns of the modern tropics but were very abundant and diverse in Paleozoic lowland swamps. This group is first recognized in the Pennsylvanian and continues to the Recent, where 90 species in six extant genera inhabit tropical areas around the world. Members of the Marattiales produce tuberous unbranched stems that can attain arborescent stature. Main axes have a dictyostele (Figure 6.11) a type of siphonostele with overlapping leaf gaps. These plants produce abundant shoot-borne roots that are large and fleshy, and often completely obscure the stem axis of the plant. They produce eusporangia that fuse into synangia borne abaxially on ultimate pinnule laminae. A modern example is Marattia; the most common ancient form is Psaronius, which is common in coal balls of the Upper Pennsylvanian.
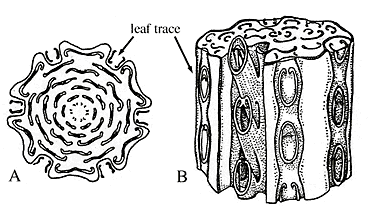
|
| Psaroniusstem (A) transverse section, (B) showing how leaf traces relates to rest of stele. |
Psaronius (Figure 6.12) was abundant in the Pennsylvanian where it may have grown up to eight meters tall and possessed a large crown of pinnately compound leaves arranged in spirals or vertical rows. The petioles of Psaronius have a characteristic "C-shaped" vascular trace that is adaxially oriented. Preserved in compression/impression, petiole scars go by the form-generic name Caulopteris. The C-shaped vascular trace is characteristic of the Marattiales and many other ferns. Psaronius had abundant shoot-borne roots that encased the stem and gave the plant a buttressed appearance. Fertile fronds bore synangia composed of eusporangia on the abaxial surface, commonly clustered near veins.
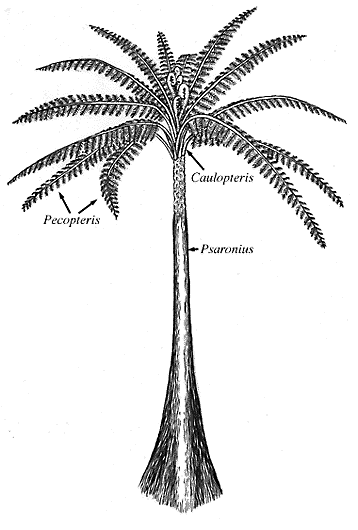
|
| Reconstruction of Psaronius. |
Like other members of this group, Psaronius has a dictyostele with endarch maturation of primary xylem. Compare longitudinal and transverse rhizome sections (refer also to Lab 1) to understand the dictyostele in three dimensions (VG 2:9). Abundant metaxylem tracheids had annular and helical thickenings. Shoot-borne roots of Psaronius have an exarch actinostele. Roots closest to the stem have a thick sclerenchymatous cortex; these reinforced roots probably helped support the tall stem of this plant, which lacked secondary xylem (VG 2:10). Fronds of Psaronius are large, up to three meters in length. They are bilaterally symmetrical and planate. Foliage is of the Pecopteris type, a foliage form genus that probably encompasses foliage from a variety of Paleozoic ferns (VG 2:11). Eusporangia are arranged in synangia on the abaxial surfaces of ultimate pinnules (VG 2:12)(VG 2:13). Sporangia have a uniform dehiscence along the midline of the sporangia. The spore genera Punctatisporites and Laevigatosporites are associated with Pecopteris foliage. Beware, however-both the spore and foliage genera are associated with stems of plants other than Psaronius.
In permineralized stems and sections of Psaronius, distinguish between the stem vascular system (VG 2:9), petiole traces, shoot-borne roots (VG 2:10), and cortex tissue within the stem. Observe the structural as well as absorptive function of shoot-borne roots. Our collection has an abundance of Pecopteris foliage from the Mazon Creek locality of Illinois (Pennsylvanian) (VG 2:11)(VG 2:12). Compare these specimens to living Marattia (VG 2:2) and Angiopteris. (VG 2:14)
Filicales. Representatives of this group, which contains most living ferns, first appeared in the Mississippian. Fossil representatives of the groups include Botryopteris, Cyathodendron, Osmunda (there are also living members of this group for you to compare), Cladophlebis, Phlebopteris, Clathropteris, Ankyropteris/Tedelea, and Tempskya. Stele morphology varies from a simple protostele to complicated dictyosteles and leaf morphology ranges from simple leaves to complex dichotomizing planar fronds. The characteristic that unites the Filicales as a monophyletic clade is the presence of leptosporangia with an annulus-an area of specialized cells involved in dehiscence (Figure 6.10) (VG 3:1)(VG 2:5). The position of the annulus is a key character used to differentiate lineages within the clade. However, sporangia are not always available for fossil ferns; and when present, determining sporangial type is often difficult. Leptosporangia typically produce only a few spores, and are most commonly homosporous. These features have been used to recognize early Filicalean ferns in the fossil record.
In sections of modern ferns, note the characteristic dictyostele in transverse and longitudinal sections of Osmunda (or Cyathea (VG 3:2). In contrast, Pteris has a protostele.
In permineralized transverse sections of Anachoropteris, a member of the Schizaeaceae from the Pennsylvanian, note the "W-shaped" petiole trace. Modern members of this family have an annulus composed of a single row of cells. In Carboniferous Anachoropteris, two rows of cells composed the dehiscence structure. Transverse sections of Botryopteris, another member of this group, show the "omega-shaped" petiole trace (VG 3:3).
Tedelea (Pennsylvanian) (Figure 6.13), which is the fertile frond of Ankyropteris, has a strongly lobed stele structure in the rachis.
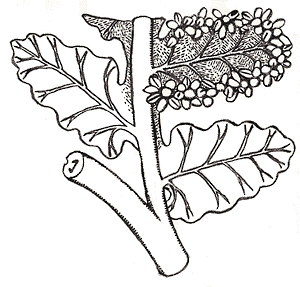
|
Part of a fertile frond, Tedelea. Note petiole trace. |
The Cretaceous genus Tempskya (Figure 6.14) is represented in our collection as permineralized stems, sections, and peels. It is the only member of the family Tempskyaceae, and a prime candidate for taxonomic revision when more material-particularly fertile fronds-are unearthed. Tempskya is known only from stem permineralizations so we have no information about foliage or sporangia. This tree-like fern has a trunk composed of many small branched axes (VG 3:4)(VG 3:5). Each axis is a siphonostele with phloem both inside and outside of the xylem ring (this feature is called an amphiphloic siphonostele). These steles and their leaf traces (be sure you can distinguish them!) are bound together in a mass of shoot-borne roots. This feature is analogous to the shoot-borne roots in Psaronius, and is also seen in some modern ferns; however, Tempskya appears to lack a dominant main stem. Examples of Tempskya stems up to 40 cm in diameter and 4.5 m tall have been recovered. Compare the growth form of Tempskya with that of Cyathodendron, the permineralized stem of a tree fern of the Cyathaceae (VG 3:6). There are living examples of Cyathaceae and its close relative, Dicksonia, growing on campus (VG 3:7).

|
| Reconstruction of Tempskya. Due to the relatively small size of the petiole and petiole traces, paleobotanists have reconstructed this plant with a stout trunk and fairly small leaves. |
Salvinia, Azolla, and Marsilea. These taxa represent a monophyletic clade of heterosporous, aquatic leptosporangiate ferns. All three genera have living representatives. While fossils of Marsilea are very rare, Salvinia and Azolla have extensive fossil records as foliage compressions (VG 3:8), and as microfossils (spores). Within the microsporangium, the microsporesare embedded in a foamy mass that is dispersed as "floats". Each float or massule is equipped with hooked projections. When released into the water column, massulae attach themselves to megasporangia and accomplish fertilization. Both massulae with their microspores and megaspores have been recovered from rocks as old as Early Cretaceous. Be on the lookout for living representatives of this group when you visit the U.C. Botanical Garden.
![[Previous Page]](../VPLimg/Back.jpeg)
|
![[Title Page]](SpheFeD/SpheFebutt.jpeg)
|
![[Glossary]](../VPLimg/Glossbutt.jpeg)
|
![[Range Chart]](../VPLimg/Rangebutt.jpeg)
|
![[Geologic Time Scale]](../VPLimg/timesbutt.jpeg) |
![[Next Page]](../VPLimg/Forward.jpeg)
|
![[authors]](../VPLimg/authorbutt.jpeg)
![[copyright]](../VPLimg/copybutt.jpeg)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}