![[Laboratory VIII - Medullosans and Cycads]](MedCycD/MedCycfont.jpeg)
Medullosa and its close relatives have been intensely studied, and as a result, a great deal is known about these strange and controversial seed plants. In part, Medullosa has attracted attention because of its avocado-sized, fleshy seeds and extremely large wing-less pollen suggesting interaction with animal dispersers and pollinators -- an evolutionary venue opened up with the advent of the seed. Medullosan organs are widely distributed in the flood plain deposits of tropical and subtropical Carboniferous, along with those of other Paleozoic seed plants such as Cordaites. The group ranged from the Mississippian to well into the Permian, and attained their greatest diversity in the Upper Pennsylvanian. They are reconstructed as trees with an overall resemblance to an old friend, Psaronius.
Medullosa is characterized by stems that appear to have a number (usually two to five) distinct vascular strands, each with a protostele that includes a number of mesarch to nearly exarch primary xylem bundles irregularly separated by parenchyma cells (Figure 8.1) (VG 1:1). Secondary manoxylic wood is produced by a bifacial vascular cambium and surrounds each vascular strand. Large rays connect parenchyma within the stele to ground tissue outside the wood. Wood development is commonly thickest toward the center of the stem.
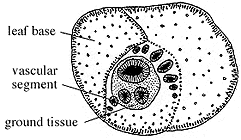
|
Figure 8.1: Transverse section of the stem genus Medullosa showing numerous vascular bundles and large leaf bases surrounding the axis. What parts of the plant are functioning as physical support for the tree-like form? |
The unusual vascular system of Medullosa has been considered "polystelic" or composed of several discrete steles by many workers. This would make Medullosa very unusual, if not unique, among seed plants, thus complicating evolutionary relationships. However, careful developmental studies (Bassinger, Rothwell and Stewart, 1974; Rothwell, 1976) have revealed that Medullosa has a form of eustele, with the unusual feature being the position of the vascular cambium. Consequently, Medullosa is really monostelic, with the vascular cambium surrounding different bundles of the eustele rather than surrounding the entire collection of primary xylem as in angiosperms. Medullosa stems have a thick, sclerenchymous cortex with abundant secretory glands and ducts. Examine and sketch cross sections of Medullosa stems (Figure 8.1) (VG 1:1).
The stem genus Sutcliffia is a close relative to Medullosa. Sutcliffia differs from Medullosa in having fewer vascular bundles, less well developed wood, and larger primary xylem. The leaf traces also have different structure.
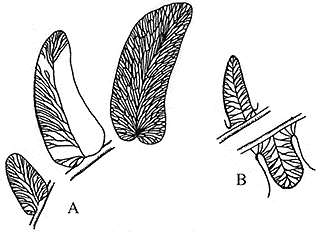
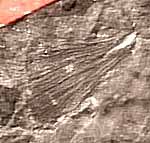
Foliage of Medullosa (here referring to the whole plant) is borne on Myeloxylon petioles, which have a plethora of leaf traces scattered among them (Figure 8.2) (VG 1:2). Fronds are usually bifurcate and have foliage described under the names Neuropteris, Alethopteris, Odontopteris, Mixoneura, and Callipteridium (among others). The Neuropteris and Alethopteris (Figure 8.2 and 8.3) types are most common, and those the you should become familiar with. Alethopteris (VG 1:3)(VG 1:4) is commonly found in organic connection with Myeloxylon petioles, but Neuropteris (VG 1:5) has only been found in association, thus its affinity with the Medullosans remains somewhat questionable. In sections of Myeloxylon, note the organization (or lack there of) of vascular traces. Compare these vascular traces with those you observed in true ferns.
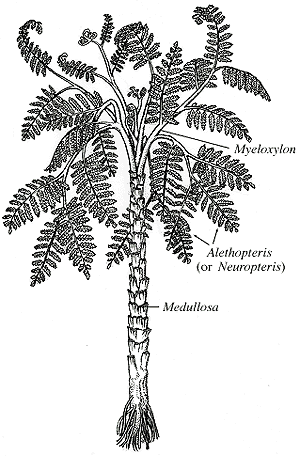
|
| Figure 8.1: Reconstruction of the whole plant, Medullosa.
|
Alethopteris and Neuropteris also differ in their internal anatomy. Alethopteris generally has small, thick pinnules with a thick epidermis and hypodermis (VG 1:3)(VG 1:4), sunken stomata, epidermal hairs, and an adaxial palisade layer with abaxial spongy mesophyll. Many Neuropteris, in contrast, has thin broad lamina (VG 1:5) with a thin epidermis and uniform, equidimensional mesophyll cells. Neuropteris species commonly lack papillae and hairs, and have dense stomatal arrays. Characters such as a palisade layer, thick epidermis, hypodermis, and hairs are common among leaves that are adapted for a sunny environment. Platy mesophyll, thin, broad lamina, and thin epidermis are more characteristic of shade leaves. These observations have led some paleobotanists to wonder whether the two types of foliage might have been borne by pteridosperms occupying different guilds within the ancient forests. Alethopteris, for example might be well adapted to high light conditions in recently disturbed environments, while Neuropteris might be well suited to the low light conditions of the forest understory. Clearly, more research is needed to confirm these speculations, but using morphological characteristics associated with certain ecological roles in the modern flora as models for interpreting the ecology of ancient plants is a productive avenue for future research in paleobotany.
| Figure 8.3: Two common foliage types associated with Medullosa stems: (A) Neuropteris and Alethopteris. |
|
The ovule of Medullosa is Trigonocarpus (when preserved in compression)(VG 1:6) and Pachytesta (when permineralized)(VG 1:7). However, these names have been sufficiently muddled so that Trigonocarpus is sometimes applied to permineralized material as well. Medullosan ovules range in size from 7 mm to 6 cm in length. The ovule is usually radiospermic, with one end of the integument drawn out into a micropyle that probably helped guide pollen to the megagametophyte within (Figure 8.4). A pollination drop mechanism may also have aided pollen capture. The nucellus is attached to the integument only at the base of the ovule. The integument of Pachytesta is three-layered with an epidermis and outer fleshy layer (sarcotesta) covering a tough fibrous (sclerotesta) and thin endotesta, which lies adjacent to the nucellus (VG 1:7). This complex structure, combined with the size of Medullosan seeds has led some to wonder about the mode of seed dispersal of these plants. However, these aspects of ancient plant ecology are notoriously difficult to evaluate scientifically.
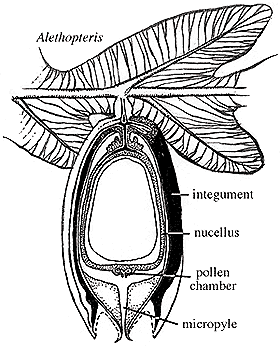
|
| Figure 8.4: Pachytesta (Trigonocarpus) ovule attached to Alethopteris foliage. The connection and orientation of the ovule is interpretive as few examples of ovules in physical connection with foliage have been found to date.
|
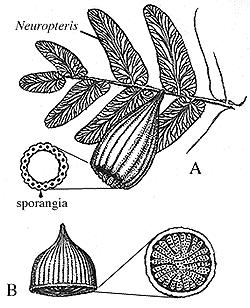
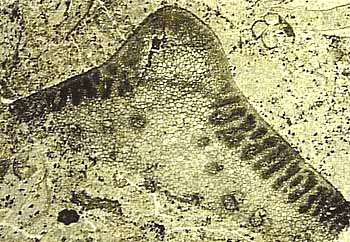
The variety of pollen organs found in association with Medullosa range from simple synangiate forms to complex, compound structures; all of which involve more or less fused clusters of elongate microsporangia. Among the simpler types Whittleseya has a uniseriate ring of sporangia within a protective cone-like structure (Figure 8.5)(VG 1:8). Dolerotheca (or Bernaultia) is one of the largest and most elaborate structures, with several in-folded synangia (Figure 8.5) (VG 1:9). Pollen of the monolete type -- called Monoletes or Schopfipollenites -- is recovered both from the pollen organs themselves and also from the micropyles of Pachytesta. The pollen is unusual because it can range from 100 mm to nearly 300 mm in length -- enormous for a pollen grain. As with the elaborate integument of Pachytesta, the large size of Monoletes pollen has led some to speculate about a pollinator interaction. Once again, this hypothesis will be almost impossible to test scientifically, but it does make an interesting story.
| Figure 8.5: Pollen organs of Medullosa: (A) Whittleseya attached to Neuropteris foliage and in transverse section. (B) Dolerotheca (Bernaultia) showing complex systems of pollen sacs. |
|
![[Previous Page]](../VPLimg/Back.jpeg) |
![[Title Page]](MedCycD/MedCycbutt.jpeg) |
![[Glossary]](../VPLimg/Glossbutt.jpeg) |
![[Range Chart]](../VPLimg/Rangebutt.jpeg) |
![[Geologic Time Scale]](../VPLimg/timesbutt.jpeg) |
![[Next Page]](../VPLimg/Forward.jpeg) |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}