![[Laboratory X - Anthophytes, Glossopterids and Others]](CupD/CupfontB.jpeg)
The occurrence of tongue-shaped leaves referred to as Glossopteris were among the first suggestions of continental drift before the breakthrough of this theory in the late 1960's. The abundant and ubiquitous presence of glossopterid leaves in Pennsylvanian to Triassic rocks of Australia, Africa, South America, Antarctica, and the Indian peninsula lead geologists to the conclusion that these continents had once formed a continuous land mass -- Gondwanaland. Moreover, the plants producing these leaves were ecosystem dominants in the Southern Hemisphere for most of this period, particularly during the Late Permian. The glossopterids are reconstructed as shrubs or large trees and probably filled a variety of ecological roles. The diversity of their reproductive structures suggests that they constituted a species-rich group, with a great number of species (over 200 described!). However, it remains uncertain exactly how much of this variation is between-taxa as opposed to within-taxon. Until 1970, when permineralized specimens of Glossopteris were discovered, commonly poorly-preserved impression/compressions were the sole basis for classification. Systematics based on leaves alone has shown to be a mission impossible (probably because there are few reliable characters). It was not until recently that ovule and pollen organs, as well as roots, were found in organic connection to Glossopteris foliage and stems. These discoveries provide evidence that Glossopteris-type of leaf was perhaps produced by several unrelated taxa. Clearly, a revision of this genus is needed!
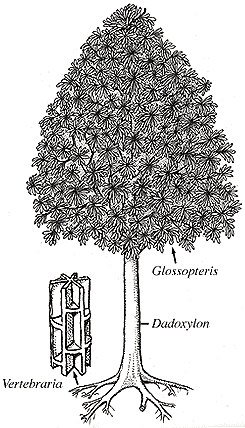
Glossopteris leaves are tongue-shaped, commonly with an entire (smooth) margin; they have a distinct mid-rib and reticulate venation adjacent to the mid-rib (VG 1:1). Some specimens exceed 30 cm in length. Glossopteris foliage may have been borne on an herb, shrub, or substantial tree (or all three). Large trunks of permineralized wood attest to the latter at least. Leaves were borne in spirals or whorls, probably on short shoots. Notice in particular the leaf venation when you examine Glossopteris leaves (Figure 10.1)(VG 1:2).
|
| Figure 10.1: Reconstruction of the whole plant, Glossopteris. Note the arrangement of secondary xylem in Vertebraria.
|
The wood of Glossopteris is classified within the form genus Dadoxylon (a.k.a. Araucarioxylon)(VG 1:3)(VG 1:4)(VG 1:5). Note that Dadoxylon wood in connection with Glossopteris foliage is indistinguishable from that produced by Cordaites and the modern conifer genus Araucaria. Commonly, Mesozoic examples of this wood type are placed in the genus Araucarioxylon but this does little to resolve to resolve the confusion or help determine the biological affinities of the wood. The message in all this may be that pycnoxylic wood anatomy -- wood that has very little ray parenchyma -- in Upper Paleozoic and Mesozoic seed plants is very conserved. This may be because directional selection on wood anatomy was fairly low or because stabilizing selection was very high (i.e., these seed plants found something that worked). What are some alternative explanations?
Vertebraria (Figure 10.1), the roots associated with the Glossopteris plant are easily recognized due to their characteristic wedge-like sections of secondary xylem radiating out from the central zone of exarch primary xylem. The wood wedges are interconnected via transverse "platforms" of wood that commonly contain a root trace. The areas between the xylem arms were probably empty during the life of the plant.
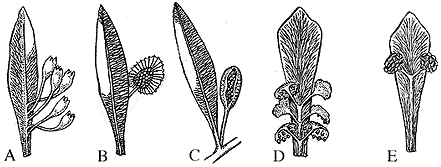
If the leaves were fairly uniform in their morphology, the reproductive structures showed substantially more variation -- even if on a single theme. Basically, both pollen and ovulate structures are borne -- separately! -- on the adaxial side of a more or less modified Glossopteris leaf (Figure 10.2). In the case of ovulate fructifications, ovules are arranged on a dorsiventral structure that has been variously called capitulum, megasporophyll, cupule (!). We have only one example of Glossopteris ovulate structures, Scutum (VG 1:6), but some other common types are illustrated in Figure 10.2: Ligettonia, Denkania, and Dictyopteridium. Pollen organs are slightly less common in the fossil record than ovulate structures, but a few types have been recognized, for example Glossotheca and Eretmonia (Figure 10.2). Eretmonia pollen sacs are born in pedicellate clusters that arise as a branch from the adaxial side of the mid vein of a modified leaf. Weird? Keep reading!
|
| Figure 10.2: Common reproductive structures associated with Glossopteris. (A) Denkania (ovulate), (B) Scutum (ovulate), (C) Dictyopteridium (ovulate), (E) Eretmonia (pollen-producing).
|
In the original description by Plumstead (1956), Scutum was interpreted as bisexual flower. Some then suggested the glossopterids as potential angiosperm progenitors. This discussion continued even after it was shown that Scutum produced only ovules. Advocates of the Glossopteris-origin theory propose the glossopterid vegetative leaf to be homologous with the angiosperms carpel, and the megasporophyll with the outer integument of an angiosperm seed, based on some peculiar specimens with abaxially borne seeds. Other paleobotanists have suggested that the glossopterids are a link between the cordaites and the gnetophytes, others ally them with the corystosperms, peltasperms and Pentoxylon, while others argue for an affinity to cycads or conifers. Once again, the question of homology becomes central to all of these arguments.
Another important Gondwanan taxon that had its heyday in the Early Permian is Gangamopteris (foliage). It is identical to Glossopteris in gross morphology, but is generally larger, has a less prominent midrib and more uniform venation. Ottokaria is the ovulate structure that has been found attached to it.
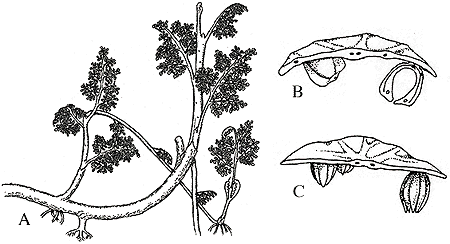
Although this genus was discovered only in the 1970's, it is probably the best known of all Paleozoic seed plants. Callistophyton, reported from the Pennsylvanian of the United States and the Upper Carboniferous of France, is reconstructed as a small shrub or vine because of its slender axes (Figure 10.3). Axes are characterized by a eustele with five bunches of vascular tissue at the edge of a substantial pith. The primary xylem is mesarch (Middle Pennsylvanian) or exarch (Upper Pennsylvanian). A bifacial vascular cambium forming manoxylic secondary wood is also present. Fronds up to 30 cm in length and of Medullopteris type are spirally arranged; branching is axial. Shoot-borne roots with exarch protosteles are common, as well as larger roots of the same type but having secondary xylem.
|
| Figure 10.3: Callistophyton. (A) Whole plant reconstuction showing vine-like growth form and leaf attachment. (B) Callospermaroin ovules attached to pinnules of Callistophyton (in transverse section). (C) Idanothekion microsporangia attached to pinnule (in cross section).
|
Callospermarion is the platyspermic seed produced by Callistophyton. It is not clear exactly where on the plant these seeds where borne. An educated guess would be on the abaxial side of pinnules, since the pollen organs, Idanothekion or Callandrium, have been found attached this way. The pollen organs are synangia, each consisting of a ring of microsporangia united at their bases around a hollow center, a common feature among so called pteridosperms (Figure 10.3). Can you recall an example from previous labs? Vesicaspora, the pollen of Callistophyton, shares many similarities with the pollen of coniferophytes. What is the significance of this? As you can imagine, a few questions remain as to the affinities of Callistophyton.
![[Previous Page]](../VPLimg/Back.jpeg) |
![[Title Page]](CupD/Cupbutt.jpeg) |
![[Glossary]](../VPLimg/Glossbutt.jpeg) |
![[Range Chart]](../VPLimg/Rangebutt.jpeg) |
![[Geologic Time Scale]](../VPLimg/timesbutt.jpeg) |
![[Next Page]](../VPLimg/Forward.jpeg) |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}