![[Laboratory IX -- <EM>Ginkgo,</EM> <EM>Cordaites,</EM> and the Conifers]](CorConD/CorConfont.jpeg)
The ginkgophytes are an ancient group that attained maximum diversity in the Jurassic. Today this group is represented by a single species, Ginkgo biloba, which is extinct in the wild, but extensively propagated as an ornamental tree. In this way, Ginkgo biloba has recaptured much of its ancient biogeographic range because it is an excellent "city tree" that grows well throughout the mid latitudes today. Ginkgo biloba could be called humankind's first effort at species preservation because it was rescued from extinction in the wild and preserved in "captivity" by the Chinese priest classes for many centuries before it came to Europe and the New World as an ornamental. Some speculate that Ginkgo biloba 's decline in the wild may be related to the extinction of a seed disperser during the Pleistocene extinctions. Ginkgo seeds are difficult to germinate, which some have suggested may be related to the requirement that they pass through the gut during dispersal. However, such a scenario is difficult if not impossible to evaluate scientifically.
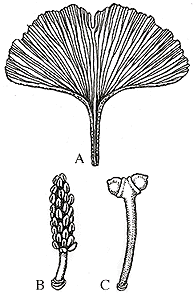
Modern Ginkgo is a tree with conifer-like architecture: A straight trunk with branches diverging at nearly right angles. Branches themselves are constructed of short shoots and long shoots (VG 1:1). In short shoots, internodes are very short, in contrast to long-internode long shoots. Secondary xylem in long shoots is pycnoxylic like that of conifers and Cordaites; wood in short shoots is rich in parenchyma, approaching the manoxylic condition of cycads. Ginkgo biloba is deciduous. Leaves develop on both long shoots and short shoots although the shape of leaves varies somewhat between long and short shoots. Leaves are generally triangular in shape with veins that bifurcate to fill space as the wedge of the leaf widens (Figure 8.1)(VG 1:1). In living Ginkgo, leaf shape is highly variable. This has led to some speculation that fossil ginkgophyte species defined on leaf shape may be over-split -- more taxa described than might have actually existed.
|
| Figure 9.1: Ginkgo biloba (A) typical leaf; (B) pollen-bearing strobilus; (C) paired ovules on stalk at the point of pollination. Ovules continue to develop and fertilization takes place after ovules fall to the ground.
|
The paired ovules of Ginkgo are borne on stalks on short shoots (Figure 8.1) (VG 1:2). In Ginkgo biloba the ovule has a three-layered integument: a fleshy outer sarcotesta, a stony inner sclerotesta, and a thin endotesta (VG 1:6). This is superficially similar to integument differentiation in Medullosa, which we also speculated might have been animal dispersed. Because of the similarity of Ginkgo biloba fruits to the fleshy fruit of conifers like Torreya, Ginkgo was originally assigned to the conifer family Taxaceae. However, subsequent character analysis has not supported this relationship. Most analyses show modern Ginkgo as a lone survivor of a lineage represented mostly by fossil plants.
Pollen is produced in strobili, which are also displayed on short shoots (Figure 8.1) (VG 1:7). Ginkgo biloba is dioecious, meaning that individual trees produce only ovules or microsporangiate strobili. This is a character in common between Ginkgo and the living cycads, which are also dioecious. Modern conifers are monoecious, meaning that both ovule-bearing and pollen cones can develop on the same plant. Since we do not know whether early ginkgophytes, cycads or even Medullosa were dioecious or monoecious, we can only guess at what node the character might have arisen.
A diversity of Permian and Mesozoic ginkgophytes have been described based on leaves, wood and some reproductive structures. Some ancient Ginkgo leaves closely resemble modern Ginkgo (VG 1:8). Others are highly dissected and resemble the typical ginkgo leaf but without tissue between the veins (VG 1:9). The oldest probable ginkgophyte dates to the Early Permian. Trichopitys lacks the short-shoot, long-shoot morphology and its ovules are born in pinnately branched systems that develop in the axils of leaves. Each branch system has up to six ovules and somewhat resembles a developmental anomaly that occurs in modern Ginkgo.
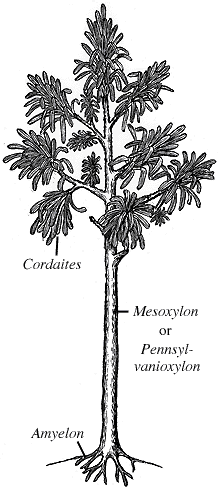
Cordaites was an important components of the Euramerican upland forests from the Pennsylvanian through the Permian. The Cordaites also contributed to coal formation as swamp plants and mangrove type trees in coastal environments. However, the greatest diversity of the Cordaites was achieved in the cool temperate forests of Angaraland, now known as the Russian-Siberian Platform.
The leaves of Cordaites are strap-shaped, commonly with parallel venation (VG 2:1). They are generally thick and leathery. In cross section, Cordaites leaves have a characteristic "I-beam" morphology created by bundles of sclerenchyma fibers above and below the vascular strands (VG 2:2) This makes these leaves very easy to recognize in coal balls.
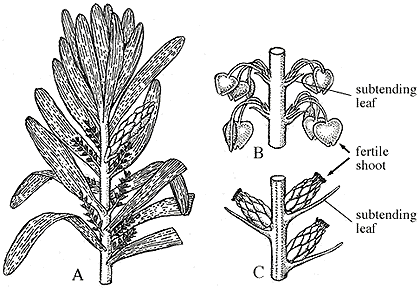
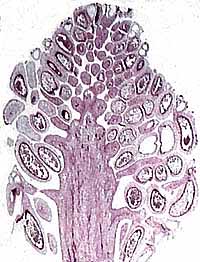
The reproductive structures of Cordaites are Cordaianthus. The form genus Cordaianthus includes both pollen- and ovule-bearing strobili. Cordaianthus consists of a primary axis with secondary shoots that lie in the axils of modified leaves (bracts) (Figure 8.2) (VG 2:3)(VG 2:4). Each secondary shoot has spirally arranged scales, most of which are sterile, but which may bear pollen sacs or ovules on short stalks. Note that this is a strobilus with possible homology to the ovule-bearing cone of conifers. Structurally preserved ovules (dispersed seeds; (VG 2:5)(VG 2:6)) are assigned to the form genera Cardiocarpus (note spelling) or Mitrospermum among others.
|
| Figure 9.2: Reproductive structures of Cordaites. (A) Foliage with fertile shoots. (B) Portion of the ovule-bearing Cordaianthus including part of the primary axis with ovulate secondary shoots bearing pendant ovules. (C) Pollen-bearing Cordaianthus including part of the primary axis with secondary fertile shoots, each of which is a compound structure with bracts subtending pollen organs.
|
The woody stems of Cordaites are placed in Mesoxylon (biseriate rays; (VG 2:7)), Cordaixylon (uniseriate rays), Dadoxylon (when found in large pieces), and Pennsylvanioxylon. All are characterized by pycnoxylic wood and a substantial pith cavity. The pith appears septate in longitudinal section due to gaps in the pith parenchyma created when cells pull apart during internode elongation (VG 2:8). Septate pith casts are called Artisia.
Roots of Cordaites are placed in the form genus Amyelon (VG 2:9). Amyelon may be either protostelic or eustelic and is surrounded by Cordaixylon-type secondary xylem. Mycorrhizal fungi have been reported in some Amyelon from Europe but the material was not sufficiently well-preserved to distinguish between a symbiotic or saprophytic role for the fungi.
|
| Figure 9.3: Reconstruction of the tree growth-form of Cordaites.
|
![[Previous Page]](../VPLimg/Back.jpeg) |
![[Title Page]](CorConD/CorConbutt.jpeg) |
![[Glossary]](../VPLimg/Glossbutt.jpeg) |
![[Range Chart]](../VPLimg/Rangebutt.jpeg) |
![[Geologic Time Scale]](../VPLimg/timesbutt.jpeg) |
![[Next Page]](../VPLimg/Forward.jpeg) |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}