The Bivalvia
Scallops, clams, mussels, and more
|
No. of described species: 10,000 |
 The Atlantic Giant Cockle (Dinocardium robustum), a heterodont bivalve. |
Bivalves are easily recognized by their two-halved shell. They can burrow into the sediment or live on the ocean floor. Some can even move around through the water by snapping their shell open and shut to swim. Not all bivalves still have a shell though; some have evolved a reduced shell or have completely lost the shell.
Throughout history, bivalves have been one of the most important marine animals to humans. They have been used for food, jewelry, decoration, even money.
Fossil record
The first occurrences of Bivalvia are found in Lower Cambrian deposits, but it is not until the Lower Ordovician that bivalve diversification, both taxonomic and ecological, explodes in the fossil record. This diversification continues unabated through the Phanerozoic, with relatively small losses at the end-Permian and end-Cretaceous extinction events. From relatively humble beginnings, they have diversified and expanded to become dominant members of most marine ecosystems.
Two extinct Cambrian bivalved groups include the Stenothecoida and the Siphonoconcha (Parkhaev 1998) — these may be nested within the Mollusca, but the absence of bilateral symmetry, enigmatic hinge structures, and shell composition places them outside of the Bivalvia as currently diagnosed.
One of the most interesting groups of bivalves, the rudists, went extinct during the Cretaceous-Tertiary mass extinction. These bivalves could be very large and often acted as the major substrate in Mesozoic reefs. Read more about rudists.
Life history & ecology
In most bivalves, the pallial cavity contains a pair of very large gills that are used to capture food particles suspended in the inhalant water current. The food is bound in mucus that is carried by cilia along food grooves on the edges of the gills to the mouth region. Here particles are sorted on the ciliated labial palps before they enter the mouth. The food then moves to the stomach, which is large and complex with sophisticated ciliary sorting mechanisms, and usually, a rotating hyaline rod which liberates enzymes into the stomach. Digestion is carried out in the large paired digestive diverticula (the stomach).
 Small, medium, and large Feathery Shipworms (Bankia setacea) at work on a submerged timber. All that can be seen of these wood-boring animals is their siphons, through which they expel undigested wood particles, the pinkish pellets that surround their holes. |
Many bivalves use their large foot to bury into the sediment on the ocean floor. They then extend a long siphon up to the surface to suck water in for filtering and breathing. You can often see these holes and the excurrent holes in mud flats at low tide. In many bivalve larvae or juveniles, a special gland, the byssal gland, can produce organic threads used for temporary attachment. In some groups, such as mussels, byssal threads permanently anchor the adults. A few groups of bivalves, such as oysters, are cemented permanently to the substrate.
Bivalves are hermaphroditic or have separate sexes. The eggs are typically small and not very yolk-rich. Fertilization is usually external, but in brooding species occurs in the mantle pallial cavity. For external fertilizers where the eggs and sperm are released into the water, the embryos often develop in the water column and go through both trochophore and veliger larval stages. Although morphologically similar to the gastropod veliger stage, phylogenetic analyses suggest that these larval stages are convergent and not homologous. Many other bivalves retain their eggs in the pallial cavity and suck in sperm with the inhalant water current. In some brooding bivalves the larvae develop in special pouches in the gills, or simply lie in the pallial cavity in others. Many brooding bivalves release their young as swimming veliger larvae while others retain them longer and release them as juveniles. Freshwater mussels (Unionoidea) have glochidial larvae that attach to fish as ectoparasites.
More on morphology
Bivalves are most easily recognized by their two shells (hence "bi-valv-ia"). Their shells are comprised of a pair of laterally-compressed hinged valves and the pallial cavity surrounds the whole body.
The bivalve shell consists of two valves that are hinged dorsally, usually with shelly interlocking teeth (the hinge), and always with a horny ligament that connects the two valves along their dorsal surfaces and acts to force the valves apart. The interior of the valves contain scars of the various muscles attached to it, in particular the (usually two, sometimes one) adductor muscles that, on contraction, close the valves. The shell can also be internal, reduced or even absent as in shipworms (Teredo, Bankia).
Bivalves typically display bilateral symmetry both in shell and anatomy, but there are significant departures from this theme in taxa such as scallops and oysters.
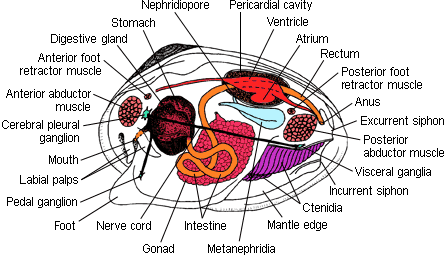 Anatomy of a clam, showing the major internal organs. |
Bivalves lack a head, radula, and jaws. Although the plesiomorphic feeding state for bivalves is probably deposit feeding utilizing long labial palps, the ctenidia provide an effective filter feeding mechanism in most taxa with numerous levels or grades of organization. In most bivalves, the mantle pallial cavity contains a pair of very large gills that are used to capture food particles suspended in the inhalant water current.
The visceral mass is primarily situated above the pallial cavity and continues ventrally into the foot. The intestine is irregularly looped and opens dorsally into the exhalant area. Also opening into this region are the paired kidneys and, when separate from the kidneys, the gonopores of the paired gonads. The heart typically lies below the center of the valves and consists of two auricles and a single ventricle that supplies both anterior and posterior aorta. The nervous system is made up of three pairs of ganglia. These innervate the musculature, mantle, viscera, ctenidia, and siphons.
The bivalve foot is modified as a powerful digging tool in many groups, while in those that live a permanently attached life (e.g., oysters), it is very reduced.
In most bivalves the two halves of the mantle are fused around the edges, with openings anteriorly for the foot and posteriorly for the exhalant opening through which the water is expelled from the pallial cavity. This exhalant also carries waste products and gametes. The inhalant opening, through which water is carried into the pallial cavity, is also posteriorly located in most bivalves, lying just below the exhalant opening. The mantle edge is also where contact is made with the external world and is, consequently where any sense organs are located. These are usually simple sensory cells but in some there are pallial eyes and/or sensory tentacles.
Systematics
Protobranchia
The protobranchs are mostly small-sized bivalves with the hinge typically composed of many small, similar teeth (taxodont condition). Notably, they include the so-called nut shells (Nuculidae). They differ from other bivalves in that their large labial palps are used in deposit feeding and the gills used only for respiration (most bivalves use their gills for filter-feeding). This group is entirely marine and the interior of the shell is nacreous in some families.
All protobranchs are shallow burrowers. One group, the Solemyidae, farm symbiotic bacteria in their gills for food and have a very reduced gut. There are about ten, mostly small-sized families in all. Many are found only in deep water.
 Protobranchs and Pteriomorphs: From left, Nucula proxima, the Atlantic Nutclam; a solemyid shell (Solemya sp.) recovered from a depth of more than 640 m in the Gulf of Mexico, off the Louisiana coast; Spondylus americanus, the Atlantic Thorny Oyster (with spines, about 11 cm long); and a group of Scorched Mussels (Brachidontes exustus) anchored to a chunk of rubble. |
Pteriomorpha
This important, entirely marine group comprises many of the most familiar (and delicious) bivalves: the scallops (Pectinidae), oysters (Ostraeidae), pearl oysters (Pteriidae), mussels (Mytilidae), and arcs (Arcidae), as well as about 18 other families. The hinge is taxodont (it has a few reduced teeth or is absent). A number of families have lost one of the adductor muscles (the monomyarian condition) and some have a nacreous shell interior.
Many pteriomorphs live as epifaunal animals being free-living, byssally attached, or cemented and have a reduced foot. The mussels seen on rocky shores in great numbers are an example of this type of lifestyle. Other pteriomorphs are shallow burrowers.
Scallops are interesting, as some species have primitive "eyes" which allow them to sense light variation. They often combine this with the ability to quickly flee from predators using jets of water squirted from between their shells.
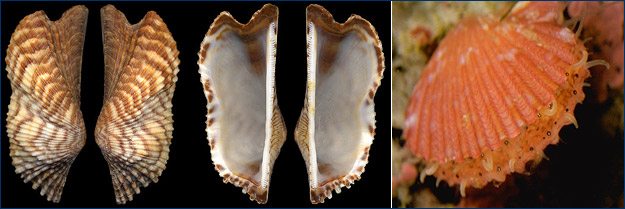 From left, a Turkey Wing (Arca zebra), an arc shell recovered off the Florida coast; a scallop (Chlamys sp.) reveals its "eyes," the black dots along the shell margins. |
Anomalodesmata
This rather diverse group includes the watering pot shells (Clavagellidae) and about a dozen other small families, some of which are found only in rather deep water. A few taxa are cemented to the substrate, but most are shallow burrowers and all are marine. One group of mostly deep-water families (collectively known as septibranchs) have the gills modified as pumping septa and feed on small crustaceans. The shells of some anomalodesmatans are nacreous and most have a simple hinge.
Rostroconchia
Rostroconches are an interesting extinct group of molluscs. It is thought that they are close relatives of modern bivalves. The larval shell was a single valve that transformed into a non-hinged, gaping bivalve shell as it grew. They probably evolved from helcionelloidean monoplacophorans in the Early Cambrian, underwent an extensive Late Cambrian and Early Ordovician radiation and survived until the Permian. Rostroconchs are thought to share common ancestry with the Bivalvia (Pojeta and Runnegar 1976; Waller 1998).
Heterodonta
This large group includes the majority of familiar burrowing bivalves: the clams, with more than 40 families including the very large family Veneridae; the cockles (Cardiidae), a family which includes the giant clams (previously Tridacnidae); mactrids or trough shells (Mactridae); and the tellins (Tellinidae). While most of the above groups are shallow burrowers, the heterodonts also include the deep-burrowing soft shelled clams (Myiidae), the ship worms (Teredinidae), and rock borers (Pholadidae). One family (Chamidae) are cemented and some members of the very diverse, mostly small-sized Galeommatoidea are commensals with a wide range of invertebrates.
 Burrowers and borers: From left, an Atlantic Mud-piddock (Barnea truncata), 4.8 cm in length; Martesia cuneiformis, the Wedge Piddock, recovered from a piece of driftwood, northeast Florida coast (1.5 cm long); and Xylophaga atlantica, the Atlantic Woodeater, recovered from trawled wood (7.5 mm long). |
The shells of heterodonts have a complex hinge comprised of relatively small numbers of different types of teeth and the shell is never nacreous. A few of their members live in fresh water (notably Corbiculidae and Sphaeriidae). The lucinids farm symbiotic bacteria in their gills which provide most of their food requirements.
Other important members of Heterodonta are the extinct rudist bivalves. These bivalves, once very important components of reef ecosystems, went extinct at the end of the Cretaceous period. Also included in the Heterodonta are the invasive zebra mussels of the superfamily Dreissenoidea.
 From left, a corbiculid heterodont, the Carolina Marshclam (Polymesoda caroliniana), 4.5 cm long. Two paleoheterodonts: a unionid freshwater Arc-mussel (Alasmidonta buckleyi) from the Altamaha River, Georgia (6.3 cm long); and the margaritiferid Western Pearlshell (Margaritifera falcata) from Lake County, Oregon (8 cm long). Note the nacreous shell interiors of these two paleoheterodonts. |
Palaeoheterodonta
The Palaeoheterodonta includes the broach shells (Trigoniidae), marine living fossils; and the freshwater mussels arranged in two superfamilies: Unionoidea (Unioniidae, Hyriidae, Margaritiferidae), and Muteloidea (Mutelidae, Mycetopodidae and Etheridae). The shell interior is often nacreous and the hinge is composed of a few, often large, teeth. All are shallow burrowers. The freshwater mussels have glochidial larva that parasitize fish.
Literature cited
Original text by Paul Bunje, UCMP. Bankia setacea and Chlamys sp. by E. Eugenia Patten, © 2001 California Academy of Sciences; all other photos courtesy of www.jaxshells.org, with Nucula proxima by Bill Frank and Xylophaga atlantica by Joel Wooster.