 |
 |
The "pelycosaurs" are members of the Synapsida, a major branch of the Amniota. Pelycosaurs are the earliest, most primitive synapsids, a group characterized by a single dermal opening in the skull permitting muscle attachment to the jaw. This means that the pelycosaurs are not reptiles, since reptiles have two such openings in their skulls. Like most other groups of synapsids, all pelycosaurs are now extinct. In fact, the only currently living synapsids are the mammals. It is believed that the pelycosaurs, like their living mammal relatives, were endothermic, which means that they maintained a constant internal body temperature. This is another characteristic that sets pelycosaurs apart from the reptiles. If this view is correct, then pelycosaurs are one of the earliest examples of endothermic animals.
In many respects, the pelycosaurs are intermediate between the reptiles and mammals, and so they have commonly been referred to as "mammal-like reptiles". The pelycosaurs indeed resemble large lizards in their overall appearance, but as we have seen, this is a misnomer since pelycosaurs are not reptiles. The name "pelycosaur" refers to a paraphyletic group of basal synapsids consisting of six separate branches: Caseasauria, Varanopseidae, Ophiacodontidae, Edaphosauridae, "Haptodonts", and Sphenacodontidae. It is currently believed that haptodonts themselves are a paraphyletic group. The Sphenacodontidae are probably the most closely related to the Therapsida (the group that includes mammals).
|
|
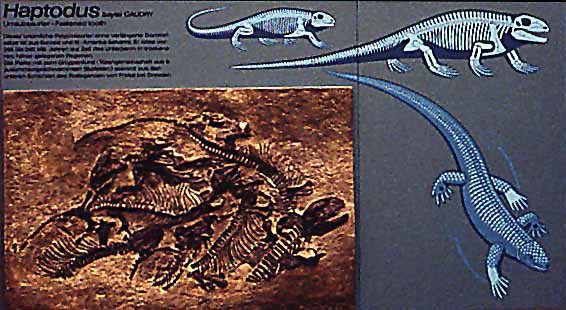
Sphenacodonts : Because the pelycosaurs are a paraphyletic group, it should not be surprising that there is considerable variety of form in the group. Among the pelycosaurs, the sphenacodonts are believed to be the group most closely related to the mammals. These include the famous "finback" Dimetrodon, shown above at left, which dominated the land fauna of the Permian but went extinct by the end of that period. The sphenacodonts also include the "haptodonts", such as Haptodus, shown above at right. (Click on either of the pictures above for a larger image).
Pelycosaurs first appeared during the upper Carboniferous (Lower Pennsylvanian) and went extinct by the end of the Permian. With the formation of the supercontinent Pangea in the Permian, continental area exceeded that of oceanic area for the first time in geological history. The result of this new global configuration was the extensive development and diversification of Permian terrestrial vertebrate fauna including pelycosaurs. Consequently, their fossils are found in multiple localities around the world. However, the majority of pelycosaur fossils have been discovered in Texas and Oklahoma, where they are among the most common fossils of the Early Permian. Much of the oil in the two states derives from the Permian period, and the search for this oil is partially responsible for the plethora of pelycosaur fossils found.
Dimetrodon is the most familiar example of a pelycosaur. A powerful carnivore, Dimetrodon was the top predator of the Permian period. With its large head and numerous large, sharp teeth, it fed on other vertebrates, including other pelycosaurs, that shared the swamps where it lived. Dimetrodon was about three-and-a-half meters (eleven feet) long and probably weighed about 100-150 kg (200-300 pounds). Four short and thick legs supported this massive body.
The physical appearance of Dimetrodon is quite distinct. While its body was long, it was relatively short. It was built like a large lizard with one important exception: a "sail". The bony sail supports rose nearly a meter vertically from its back and were connected by highly vascularized skin. The most popular conjecture regarding the function of the sail is thermoregulation. Such a sail would allow a large area for acquiring or releasing heat. Studies have shown that the area of the sail is proportional to the volume of the Dimetrodon body, as would be expected if the sail were important for heat exchange. While many tetrapods in many lineages have evolved similar sail-like structures, Dimetrodon is the best known so the majority of work done exploring the function of the sail has centered on this pelycosaur.
However, It must be noted that not all pelycosaurs had sails. Still, most pelycosaurs were similar to Dimetrodon in that they also were carnivores with large, powerful jaws, and two types of teeth: sharp canines and shearing teeth. Two major exceptions to this meat diet were the Caseidae and the Edaphosauridae, who were herbivores. While the Caseids were far more diverse, the most familiar of the herbivorous pelycosaurs is another sail-back, Edaphosaurus. Edaphosaurus had a smaller skull than Dimetrodon, and had large peg-like teeth for grinding and crushing plants. This adaptation allowed pelycosaurs to flourish, and by the beginning of the Permian, pelycosaur genera account for 70 percent of all the known amniotes, outnumbering the reptiles.
 |
 |
Edaphosaurus : Pelycosaurs include a variety of forms and lifestyles. Not all of them were carnivores; some, like Edaphosaurus, were herbivores. In the image above at left, you can see that there are no nasty, pointy teeth filling this pelycosaur's jaw. You may also see the additional knobs sticking out from the dorsal fin bones, which are unique to the Edaphosauridae. At right, you can see a fully mounted skeleton from the Permian of Texas. The long tail and sprawling posture are characteristic of the pelycosaurs. These images were taken at the Field Museum of Natural History in Chicago. (Click on either of the pictures above for a larger image).
Pelycosaurs are an important lineage preserved in the fossil record. They are the earliest known synapsids, the first to evolve specializations that would play an important role in the rise to mammals. They are thus a transitional group in both form and functions. Schad (1994) presents two general rules concerning transitional areas. Firstly, no intermediate form represents a totally intermediate character. This is to say that no evolutionary condition has been found in any taxon where all characteristics are present of both taxa that the condition is intermediate to. Secondly, evolutionary transformation predominates in the extremities, and only later affects the axial system. Bearing in mind these principles, we can begin to understand the development of mammals from their pelycosaur roots.
The pelycosaurs exhibit the first substantial progress of crawling to running. This evolution in the extremities required a modification of the metabolism in the muscular system to provide the energy reuired for more strenuous activity. The resulting change in the axial system brought about endothermy. Supporting this idea is the fact that as later pelycosaurs and later synapsids evolved, the surface area of sail to body mass ratio decreased. This shows the trend of reduced need for outside thermoregulation, which would require an increased use of endothermy, an important characteristic today separating the reptiles and mammals.
Visit the Synapsida pages at the Tree of Life, which includes a full discussion of the autapomorphies of the main clades of synapsids, with a focus on the basal (pelycosaur) groups. Or browse Jack Conrad's pages on Synapsida for additional information.
Sources:
Bennet, S. Christopher. 1996. Aerodynamics and thermoregulatory function
of the dorsal sail of Edaphosaurus. Paleobiology 22: 496-506.
Carroll, R. L. 1988. Vertebrate Paleontology and Evolution. New York: W. H. Freeman.
Florides, G. A., L. C. Wrobel, S. A. Kalogirou, S. A. Tassou. 1999. A thermal model for reptiles and pelycosaurs. Journal of Thermal Biology 24: 1-3.
Hillenius, Willem J. 1994. Turbinates in the Therapsids: Evidence for Late Permian origins of mammalian endothermy. Evolution 48: 207-229.
Hopson, James A. 1994. Synapsid evolution and the radiation of non-eutherian mamals. Pp 190-219 in D. R. Prothero & R. M. Schoch (eds.) Major Features of Vertebrate Evolution. Short Courses in Paleontology 7.
Schad, Wolfgang. 1994. Heterochronical patterns of evolution in the transitional stages of vertebrate classes. Acta Biotheoretica 41(4): 383-389.
Sidor, C. A. & J. A. Hopson. 1998. Ghost lineages and "mammalness": Assessing the temporal pattern of character acquisition in the Synapsida. Paleobiology 24: 254-273.

