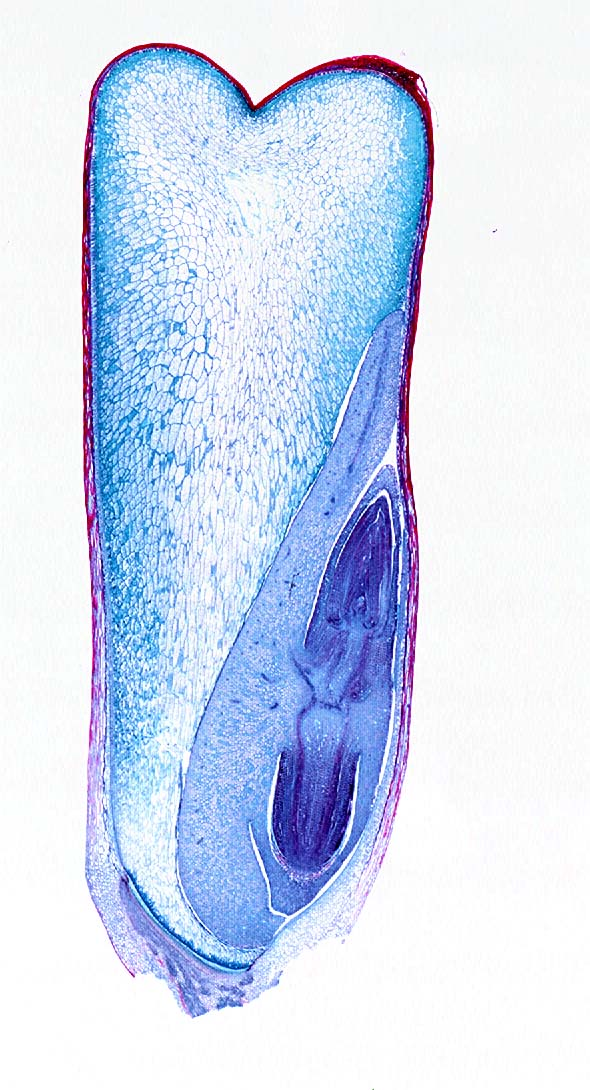
The three characters which are most closely congruent with the usual definition of monocots are the embryo morphology, stem vascular structure, and the plastid protein inclusions. The embryo of each seed plant possesses one or more leaf-like structures called cotyledons, which absorb nutrients and may function as the first leaves in some plants. Monocots have a single such cotyledon, while other flowering plants usually have two. The exception to this is the water lily Nymphaea, which has what appears to be a single cotyledon with two lobes, but which has usually been interpreted as two fused cotyledons. Since the water lilies are quite closely related to the monocots, this may represent a transitional form. The image below at right shows a monocot embryo with cotyledon.

| 
| 
|
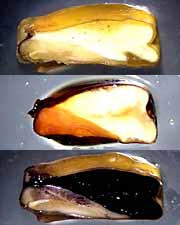
Monocot characteristics : At far left, a cross section through the stem of a corn stalk. The small dark circles visible in the stem are strands of vascular tissue; imagine you are looking down on the end of pipes that run up and down through the corn stem. The vascular strands in monocots are not arranged in a neat ring, as they are in dicots, but are scattered within the stem. At right, images of a corn kernel. The kernel is the fruit produced from the corn flower, and each kernel contains an embryo that may develop into a new corn plant. The embryo has only one cotyledon, which is a part of the embryo used to absorb the nutrients stored in the endosperm, a food reserve stored for the young plant. At center above, are corn kernels stained to show chemical composition. The top central image is an unstained kernel sliced open. Below it is a kernel stained for protein (orange), showing that the protein is concentrated in the embryo and its cotyledon. Below that is a kernel stained for starch (black), showing that the endosperm is mostly starch. Compare with the image at right.
The second character common to all monocots is the atactostele. This refers to the arrangement of vascular strands in the stem, which in the case of monocots are scattered in the stem, though more densely packed near the periphery of the stem. This feature is not unique to monocots, but is found also in the paleoherbs and certain magnolia-like dicots.
The third feature common to all monocots is the structure of certain plastid inclusions. All plant cells contain plastids, some of which become photosynthetic, but those which are found in the phloem tissue do not do so. When examined in properly prepared material, the plastids in the phloem are shown to contain crystalloid proteins, the shape of which is distinctive for different major groups of plants. In the monocots and a few paleoherbs, these crystalloids have a triangular shape, unlike all other flowering plants. This is an obscure character, but it is reliable.

 A number of monocots have developed tree-like forms in which the leaves
attach only at the base of the plant. These include the massive agaves as well
as bananas and the so-called traveller's palm. In these cases, the leaves may
occur only near the base of the plant, forming a rosette, or they may
wrap around the stem, lending it support, as in bananas. Some of the rosette
forms have evolved succulent leaves, which are fleshy and store water.
These include the agaves, aloes, and many bromeliads.
A number of monocots have developed tree-like forms in which the leaves
attach only at the base of the plant. These include the massive agaves as well
as bananas and the so-called traveller's palm. In these cases, the leaves may
occur only near the base of the plant, forming a rosette, or they may
wrap around the stem, lending it support, as in bananas. Some of the rosette
forms have evolved succulent leaves, which are fleshy and store water.
These include the agaves, aloes, and many bromeliads.
Two other morphological forms of some importance in the monocots are those herbs which grow from bulbs, and those which grow as vines. A wide range of monocots produce bulbs of modified leaves which store food over the winter. An onion is an examle of one such plant, and the layers of an onion are in fact these modified leaves. Tulips, gladiolus, and lilies are other monocots which produce bulbs, as is Fritillaria, whose bulbs are shown at right. Vines are relatively rare among monocots, but three of these are commonly encountered: Dioscorea (Chinese yam), Smilax (greenbrier), and Tillandsia (Spanish "moss"). There are also a number of arums which grow as vines, such as Philodendron.