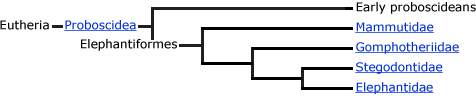
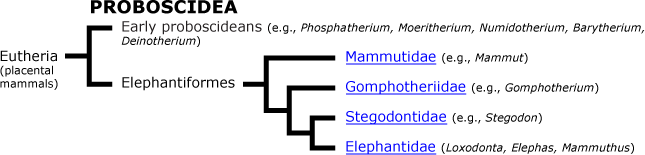
The Proboscidea
Elephants, mammoths, mastodons
 The African Elephant, Loxodonta africana. |
Today there are two living species of Proboscidea: the African elephant Loxodonta africanus and the Asian elephant Elephas maximus. However, in the past, there were many members of the order.
Early relatives
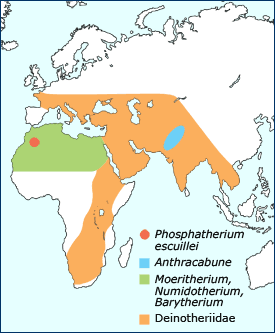 Ancestral proboscidean distribution. |
Other ancestral proboscideans include Numidotherium, Barytherium, and Deinotherium. Remains of these early proboscideans were recovered in northern Africa along what was once the southern shore of the Tethys Sea during the Oligocene. Numidotherium stood about 1.5 meters tall and had a trunk about the length of a tapir's. Many remains of this early proboscidean have come from middle Eocene deposits in Algeria. There were two species of Barytherium. One species was large, weighing about 3–4 tons and standing 2.5–3 meters high. The second species was smaller, about the size of Moeritherium. Barytherium lived from the late Eocene to the early Oligocene. Deinotherium had a fully functioning trunk and downward recurving tusks in the lower jaw, a trait present in no other proboscidean. It is thought that they used the tusks for manipulating their short trunk or for mate recognition, but not for digging. They had brachydont cheek teeth ideal for shearing soft foliage. Deinotheres roamed the planet for 20 million years, living in Africa, Europe and Asia beginning in the middle Miocene; they persisted in Eurasia until the Pliocene and in Africa until just over one million years ago.
It is hypothesized that anthracobunids (Anthracobune) belong to the proboscidean order, or at the very least, are closely related to it. They lived in southern Asia during the early Eocene. If they do belong to Proboscidea, their presence in Asia broadens the distribution of early Proboscidea.
|
Additional UCMP resources
Other resources
|
||
|
Sources
|
||
Original Proboscidea page was by Brian R. Speer, 7/8/1997; Dave Smith made minor design changes and link updates, 7/15/2005; major new content researched and written by Kaitlin Maguire, 11/2010; elephant photo by Gerald and Buff Corsi, © 2002 California Academy of Sciences