How Did It All Begin? The Self-Assembly of Organic Molecules and the Origin of Cellular Life
by David W. DeamerFrom Evolution: Investigating the Evidence, Paleontological Society Special Publication Volume 9, 1999. By permission of the Paleontological Society.
Introduction
Movies are the myths of late-20th century western culture. Because of the power of films like ET to capture our imagination, we are more likely than past generations to accept the possibility that life exists elsewhere in our galaxy. Such a myth can be used to sketch the main themes of this chapter, which concern the origin of life on the Earth.
Imagine that 4 billion years ago, intelligent beings evolved on an Earth-like planet in the solar system of a neighboring star. After ten million years of evolution, they have solved the problems of interstellar travel and aging. Virtually immortal family groups set out to explore the galaxy and almost immediately discover a solar system associated with a nearby star only 80 light years away from their home planet. They find that the third planet is rich in the primary elements of life — carbon, hydrogen, oxygen and nitrogen — which are present in the atmosphere in the form of carbon dioxide (CO2), molecular nitrogen (N2) and water vapor (H2O). They decide to spend a few centuries studying this planet, which they consider to be a possible model of their own primordial world as it was four billion years in their past.
They learn that the planet gained most of its mass through a process called accretion, in which gravitational attraction causes dust particles to gather first into small asteroid-sized planetesimals, which then undergo immensely energetic collisions to form ever larger planetary bodies. A single moon looms in the sky, the result of one such collision during the final stages of accretion. The planet and its moon were molten after the collision, reaching temperatures of volcanic lava which degraded all organic compounds, leaving only their elements in the form of volatile gases. Additional carbon dioxide and water vapor continued to pour into the atmosphere through volcanoes that allowed outgassing from the planet's interior. The planetary and lunar crusts soon cooled by radiating heat into outer space, so that the moon's hardened surface began to preserve the accumulating record of impacts over the several hundred million years that followed the moon-forming collision event.
Now the global temperature has fallen to the point that water has condensed into vast, shallow oceans that virtually cover the planetary surface, with lava and pumice islands rising from undersea volcanic regions. Because of the very high atmospheric pressure of carbon dioxide, the temperature of the oceans is still well above the boiling point of water on their home planet. The planet is devoid of life, sterilized by the heat energy released by the original collision and later giant impacts. But the impacts have largely ceased, and the temperature continues to drop to the point at which organic carbon compounds remain stable for days, years, then decades. Some of these compounds are continuously synthesized at the planet's surface by a variety of chemical reactions, often using light as an energy source, while others are delivered by the infall of microscopic dust particles that is still adding the last few kilometers to the planet's diameter. As a result, organic substances begin to accumulate in the shallow seas, then become more concentrated by evaporation of tide pools and adsorption to mineral surfaces in submarine geothermal regions. The visitors take samples of the foamy material in the tide pools and examine it with their powerful microscopes. There is nothing alive, but they are surprised to find that some of the organic compounds have spontaneously aggregated into a variety of structures, including long molecules called polymers and microscopic bubbles. What could this mean?
The visitors fly away at faster than light velocities to visit other stars and solar systems, but return several hundred million years later to see what is happening on the third planet. (Because they can travel at supraluminal velocity, only a few years have passed in ship time.) This time they are astonished to discover that something remarkable has occurred. Instead of a sterile environment, the seas now teem with microbial life that has already begun to change the planet's surface and atmosphere. Comets and meteorites still bombard the planet, but now tens of millions of years separate the giant impacts and their energies hardly disturb the robust microbial life that has invaded every available niche.
Could their own versions of bacteria have somehow infected the planet during their last visit? A bit worried, the visitors again take samples and determine that the new life has an entirely different genetic code than their own, so it is not something they accidentally released. Apparently the spontaneous self-assembly processes they observed earlier produced enormous numbers of microscopic structures, a few of which increasingly were able to use the energy and nutrients available in the environment to reproduce their structures. But because several hundred million years have passed, the visitors missed the exact point in time at which this occurred! Frustrated, they depart for further exploration, hoping to find other primitive planets where they might have a chance to observe the origin of life. Another 3.5 billion years pass, and they return once again to see what has transpired. By this time they have discovered that planets with liquid water and life are common in the galaxy, so they are not surprised to find that primitive yet intelligent organisms now inhabit a place called California. When their space ship is observed they must quickly depart, accidentally leaving behind one of their younger family members...
Evidence related to the origin of life
Just kidding about that last part. But even the mythical visitors were astonished, as we should be, that the life process can begin spontaneously on an utterly sterile planet, as long as the environment has liquid water, organic carbon compounds, and energy. How does life begin? This is one of the remaining great questions facing science, a question so daunting that it has tended to inhibit serious scientific inquiry. Even Darwin once noted that "It is as absurd to think about the origin of life as it is to think about the origin of matter." But now, 150 years after Darwin expressed his concern, we do have a pretty good idea about the origin of matter. And we understand living cells in remarkable detail, even to the point that we have begun to manipulate the genetic blueprint of life and will soon know the entire sequence of 3 billion nucleotide bases in the human genome. But we still don't know how the life process began on the early Earth.
On the other hand, we can make some informed guesses, something not possible fifty years ago when the first research on life's origins began. The main point to be made here is that certain kinds of molecules have physical and chemical properties that allow them to self-assemble into orderly structures, and these are the molecules used by living cells. The self-assembly process seems to defy our intuitive expectation from the laws of physics that everything on average becomes more disordered (i.e., entropy increases). For instance, it is easy to calculate that it would be impossible for a specific protein ever to be produced by chance in the lifetime of the universe, and therefore conclude that a supreme being designed all proteins. However, one can make the same argument with a soap bubble. A bubble can't exist, according to the laws of chance, yet they are a common occurrence.
A number of recent scientific advances have provided a more coherent picture of the events leading up to the origin of life. This integrated vision has given rise to a new field called astrobiology, defined as the investigation of life in the universe, and takes into account our new understanding that life on the Earth is part of a universal process. The following question-answer format indicates the things we know with a fair degree of confidence, that now provide a framework for developing and testing hypotheses related to the origin of life.
Where does matter come from?
All atoms heavier that hydrogen and helium, including the elements important in living systems (carbon, oxygen, nitrogen, phosphorus and sulfur) are produced are produced in stars by nuclear fusion reactions. The atoms are then blown out into interstellar space toward the end of a star's lifetime when the star explodes as a nova or, more rarely, a supernova. The atoms then form molecules and dust particles and gather into the enormous clouds that have been visualized by the Hubble Telescope in extraordinary detail. The dust particles, composed largely of silicate minerals, are called interstellar grains. The grains are coated thin layers of ice and frozen gases like carbon dioxide, carbon monoxide, ammonia and methanol, as well as a variety of more complex organic compounds. The last point is among the most significant new discoveries about the interstellar medium. That is, organic compounds composed of carbon and the other biogenic elements are not limited to the Earth and its neighboring planets in our solar system, but are present wherever stardust gathers into interstellar clouds. We live in an organic universe.
Where do stars and solar systems form?
These clouds are the birthplaces of new generations of stars. During star formation, gravity causes portions of the clouds to form rotating disks with the star at the center. Kilometer-sized objects called planetesimals are produced by gravity-driven accretion of dust within the disk, and the planetesimals undergo increasingly violent collisions to produce larger planets. Our own moon is the result of such a collision between the primitive Earth and another planet the size of Mars or larger. We now have convincing evidence of a dozen Jupiter-sized planets around other stars, and the search is on for Earth-like planets.
What are sources of organic matter on planets?
During late accretion of Earth-like planets, organic compounds and water are delivered to planetary surfaces by comets and meteors. Surprisingly, a fraction of the organic compounds are able to stay intact during their arrival in the Earth's atmosphere. We can still see the delivery of organic compounds to the Earth's surface in the form of carbonaceous meteorites, both large and microscopic. It is likely that other organic compounds were synthesized in the Earth's atmosphere, because experiments have been conducted that reproduce the process under simulated prebiotic conditions. However, because the Earth's atmosphere now contains highly reactive oxygen as a result of photosynthetic activity, if organic compounds were synthesized by abiotic processes today they would be rapidly degraded by oxidation.
What is meant by self-assembly?
Certain organic compounds have the capacity to react with each other to form more complex molecules such as hydrocarbons, amino acids and simple sugars. Some of these can spontaneously self-assemble into membrane structures, and others can polymerize into molecules similar to proteins and nucleic acids. These in turn form larger self-assembled structures such as the double helix of DNA. Because there were no genes or enzymes available on the prebiotic Earth to direct the metabolism and reproduction characteristic of living organisms, the first forms of life must have been produced through a spontaneous self-assembly process.
How did catalytic activity become incorporated into the earliest forms of life?
Most life on the Earth today depends on protein catalysts called enzymes, which like all catalysts can greatly increase the rates at which reactions proceed. Catalysts are essential to life, but what were the first catalysts? They were probably not proteins, because protein synthesis requires DNA and other chemicals combined in a process much too complex to occurred spontaneously. However, we now know that certain kinds of ribonucleic acid (RNA) also have catalytic activity, behaving like protein enzymes. These are called ribozymes, and they considerably simplify our thinking about the beginning of life. We no longer need to consider that entire cells complete with DNA, RNA, ribosomes and protein synthesis somehow appeared. Instead, we can conceive of a primitive RNA system that could grow, reproduce and evolve, showing all the properties we associate with the living state.
When did life begin?
Evidence in the form of microscopic fossils has convincingly shown that bacterial life was abundant in shallow seas about 3.5 billion years ago. Other evidence from measurements of stable isotopes of carbon suggest that even simpler forms of life existed 3.8 billion years ago. If so, it follows that life can begin in as little time as 100 million years, since the Earth's surface was probably still too hot for any conceivable form of life over 4.0 billion years ago.
Where did life begin?
The origin of life must have occurred in an environment where the temperature was low enough to permit liquid water to exist. Some examples include tide pools or sub-surface sites similar to a hydrothermal vent or a geothermal hot spring. Liquid water is required for life as we know it, since only water can provide a universal medium in which self-assembly processes and metabolism can occur. The temperature of the site was likely to have been higher than prevails on the Earth today. We now know that microbial populations can inhabit environments once thought impossible, where temperatures range up to the boiling point of water. This greatly expands the possible range of sites for the origin of life, and suggests that living microorganisms could be present in deep hydrothermal regions under the Martian crust, or even on Europa, which is now believed to have ice-covered oceans of liquid water.
The Prebiotic Earth
Up until the discovery that microbial life can exist near submarine hydrothermal vents at temperatures above 100° C, the consensus has been that life surely began at the Earth's surface. The reasons are easily understood. It seems likely that organic compounds in the prebiotic environment would accumulate as water-soluble compounds and surface films in the early ocean. Fluctuating environments such as tide pools would then provide a mechanism for concentrating the dilute solutions, and as the concentrated mixtures of organic compounds were dried and heated, an ongoing synthesis of polymeric material would take place (Fox and Harada, 1958; Usher, 1976).
Recent advances in our understanding of the primitive Earth have forced us to rethink this assumption. For instance, the lunar cratering record suggests that the Earth was subjected to giant impacts of comets and asteroid-sized objects about the time that the first living organisms appeared. The magnitude of energy associated with such events would vaporize some or all of the early ocean, virtually sterilizing the upper portion of the Earth's surface. The origin of life on the surface could only have occurred after the last such event.
The discovery of hydrothermal vents provided an alternative site for life's origin. Shortly after their discovery Corliss et al., (1981) and Baross and Hoffman (1985) proposed vents as a potential site for the first life on the Earth. More recently, Stevens et al., (1995) reported that hydrothermal regions surprisingly deep in the Earth's crust also have extensive microbial populations. Both hydrothermal vents and deep geothermal regions may have provided a refuge from giant impacts that sterilized the surface of the early Earth. This idea is supported by evidence from ribosomal RNA sequences which strongly suggests that the last common ancestor of all life on Earth was likely to have been a thermophilic microorganism (Woese, 1987; Pace, 1991).
A third alternative site was proposed by Bada et al., (1994) who noted that current models of solar evolution predict a young sun 20–30% less luminous than today's sun. Unless there was significant greenhouse warming, the early oceans would freeze to form an ice sheet that would be periodically thawed by impact events. It is well known that organic compounds such as amino acids have finite lifetimes in solution, and their stability decreases markedly as the temperature increases. At the highest temperatures associated with hydrothermal vents, amino acids cannot survive at all (Bada et al., 1994). A global ice cover and colder temperatures would afford significant protection against thermal degradation. It also seems likely that the concentrated mixture of solutes available during thaws could undergo a burst of chemical reactions leading to more complex molecules, a few of which could be on the evolutionary pathway to life.
To summarize, three alternative sites have now been proposed for the accumulation of organic compounds pertinent to the origin of life. Two of the alternatives place the site at the Earth's surface, the main variable being the temperature of the site. The third alternative places the site at a subsurface region associated with relatively high temperatures. The surface sites have access to light energy, chemical energy and to concentrating mechanisms, while the subsurface site has access to chemical energy in the form of certain mineral surfaces (pyrite and clays), dissolved gases like hydrogen and methane, and solutes such as ferrous iron.
The three alternatives do not represent hypotheses per se, but are more in the form of conjectures. That is, they can never be tested by direct experiment, because the origin of life occurred in an unknown environment over 3.5 billion years ago in conditions that cannot be reproduced with certainty today.
How did organic compounds appear on the early Earth?
The classic experiments of Miller (1953) showed that impressive yields of certain amino acids can be obtained when a mixture of gases (hydrogen, methane, ammonia and water vapor) is exposed to an electrical discharge. This discovery represented a major breakthrough, since amino acids are the monomers that compose all proteins. The mixture was assumed to be a simulation of the original terrestrial atmosphere which, by analogy with the outer planets, would have contained hydrogen, methane, ammonia and water vapor. At sufficiently high energy fluxes, such reducing systems of gases generate hydrogen cyanide (HCN) and formaldehyde (HCHO), which in turn react to produce amino acids. Cyanide and formaldehyde are now considered to be key reactants in simulations of prebiotic chemical pathways (Ferris and Hagan, 1984).
The possibility that organic material could readily be synthesized in prebiotic conditions was supported when it was discovered that carbonaceous meteorites also contained amino acids, hydrocarbons, and even traces of purines, one of the basic components of nucleic acids (Kvenvolden et al., 1970). If carbonaceous meteorites represent samples of the primitive solar system, it is reasonable to assume that similar synthetic chemical reactions may have occurred on the Earth's surface. With organic monomers available in reasonable concentrations on a global scale, it was not difficult to imagine that self-assembling systems of polymerized macromolecules would at some point assume the properties of the living state: the ability to exist as a membrane bounded entity, or cell, the capacity to use energy and nutrients to grow by directed polymerization processes, and the ability to divide in some way to produce another generation.
However, more recent results indicate that the early atmosphere was composed of carbon dioxide and nitrogen rather than a mixture of reduced gases (Holland, 1984; Kasting and Ackerman, 1986). Carbon dioxide does not support the rich array of synthetic pathways leading to possible monomers, and alternative sources of organic material are being considered. One possibility is that extraterrestrial infall in the form of comets and meteorites provided significant amounts of organic carbon to the Earth's surface (Oro, 1961; Anders, 1989; Chyba and Sagan, 1992).
An extraterrestrial source of organic compounds is surprising, but there is no doubt that all of the biogenic elements on the Earth (carbon, hydrogen, oxygen, nitrogen, phosphorus and sulfur) had an extraterrestrial origin simply because they were delivered by the Earth's accretion process. The question is how much chemical processing occurred before the elements were incorporated into primitive forms of life. Most likely both synthesis and infall were involved to some degree. For instance, amino acids are present only in trace amounts in carbonaceous meteorites, and furthermore are relatively unstable in water. It follows that amino acids and other water soluble organic compounds probably were synthesized continuously at the Earth's surface. On the other hand, hydrocarbon derivatives are relatively stable and compose several percent of the mass of carbonaceous meteorites, yet are not major products of Miller-Urey type reactions. It may be that hydrocarbons required by early life were primarily delivered with meteoritic infall, rather that being synthesized by terrestrial chemistry.
Estimates of accumulated organic compounds during a hundred million year late accretion period ending around 3.9 billion years ago are in the range of 1016–1018 kgs. This is less than the total organic carbon stored as oil shales, coal and other fossil deposits on the Earth (1021 kg) which represents carbon dioxide reduced to organic compounds by photosynthetic processes. On the other hand, it is several orders of magnitude greater than the organic carbon now in the biosphere, estimated to be 6 X 1014 kg. To put this value into perspective, if 6 X 1014 kg of the organic substances in the total biomass today were spread over the entire surface of the Earth, including the oceans, it would form a layer about 1 mm thick. For comparison, the amount of organic substances delivered by infall would form a layer ranging from 1.6 cm to 1.6 m thick. Although this amount of material would represent a significant source of organic carbon in the prebiotic environment if it all survived and accumulated, most of the cometary and meteoritic infall surviving atmospheric entry would presumably fall into oceans. A major fraction of the organic content would be buried as sediment, and a smaller fraction would be released into the marine environment over long time intervals. Water soluble compounds would dissolve to form a very dilute solution of organic solutes, while longer chain hydrocarbons and their derivatives would accumulate at the ocean surface to form a thin film at the air-water interface. Such films would likely become concentrated at intertidal zones by the same mechanism that forms sea foam from active organic compounds today. It follows that probable sites for the physical and chemical processes leading to the origin of living cells would be tide pools or geothermal region in which hydrocarbon derivatives accumulated and were mixed with water-soluble organics during cyclic drying and rehydration processes.
Self-assembly processes: soap bubbles and membranes
All life today has units that we call cells. Did life arise from pre-existing cellular structures, or did cellular life develop only at a later evolutionary stage? As noted earlier, there were no large molecules like nucleic acids and proteins available on the prebiotic Earth to direct the assembly processes characteristic of life, so the first forms of life must have arisen through self-assembly processes. Familiar examples of such processes today include the formation of bubbles from soap molecules, or the foam that accumulates in tide pools. The main point to be made in this section is that certain kinds of organic compounds called amphiphilic molecules are able to self-assemble into microscopic bubble-like structures. Such structures form spontaneously, and perhaps provided the original membrane-bounded environment required for cellular life to begin.
We will first outline the biophysical principles that govern self-assembly processes and indicate how they can be used to investigate the origin of cellular systems.
1. Bilayers assemble from of variety of amphiphilic compounds
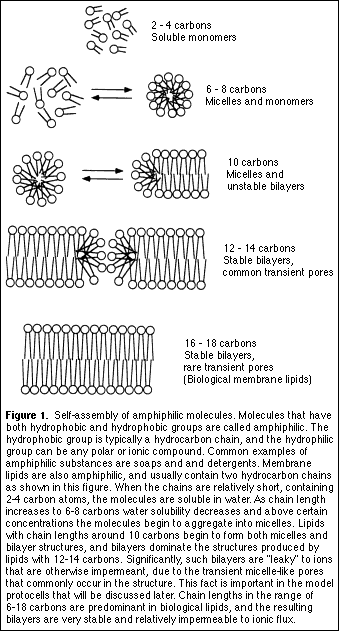
Although contemporary cell membranes incorporate phospholipids as the primary component of the bilayer, it is not necessary to think that such complex molecules were required for early cellular life. In fact, simpler amphiphilic molecules can also assemble into bilayer membranes, even single-chain amphiphiles such as soap molecules. It seems likely that primitive cells incorporated lipid-like molecules from the environment as a nutrient, rather than undertaking the much more complex process of synthesizing complex lipids by an enzyme-catalyzed process.
2. Bilayer permeability strongly depends on chain length of the amphiphilic molecules
We tend to think of the lipid bilayer as being a nearly impenetrable barrier to ionic solutes such as salt (sodium chloride) and other large, polar molecules like amino acids. But then how did early cellular life function in the absence of highly evolved transport enzymes that translocate ionic nutrients and metabolites across the bilayer barrier? It is true that lipid bilayers of contemporary cell membranes present a significant permeability barrier that is necessary for normal cell functions, particularly those related to bioenergetics of ion transport and chemiosmotic ATP synthesis. However, recent results show that shortening lipid chain length from 18 to 14 carbons increases the permeation of ionic solutes by several orders of magnitude. This level of permeability is sufficient to encapsulate large molecules such as proteins and polynucleotides, yet still allow external substrate to reach an encapsulated enzyme. It follows that early cell membranes could have been composed of shorter chain lipids that provided access to nutrients for macromolecules undergoing growth and replication in an encapsulated microenvironment.
3. Macromolecules can be encapsulated in bilayer vesicles under simulated prebiotic conditions
A third conceptual problem has been to imagine how lipid bilayers could capture macromolecules in the first place, given that the bilayer must present a nearly impenetrable barrier if the macromolecules are to be maintained within the membrane bounded volume. We will describe how a mixture of lipid and protein or nucleic acids can undergo drying and wetting cycles that simulate tide pools. Under these conditions, the macromolecules are readily captured in membrane-bounded vesicles.
4. Lipid bilayers grow by addition of amphiphilic compounds present in the environment It is not sufficient for a primitive cell to replicate its macromolecular components unless the boundary membrane can increase in area to accommodate the internal growth. Recent experimental results from liposome model systems demonstrate that such growth through addition of amphiphilic molecules can in fact occur.
Self-assembly processes in prebiotic organic mixtures
The first suggestion that membranes played a role in the origin of life was in J.B.S. Haldane's prescient note in The Rationalist Annual (1926). Haldane wrote that "The cell consists of numerous half-living chemical molecules suspended in water and enclosed in an oily film. When the whole sea was a vast chemical laboratory the conditions for the formation of such films must have been relatively favorable . . ." Goldacre (1958) proposed that the first membranes could have been produced by wave action disturbing films of lipid-like surfactants. The first experimental approaches to this question were undertaken by Hargreaves et al., (1977) and Oro and co-workers (1978).
What physical properties are required if a molecule is to become incorporated into a stable bilayer? All bilayer-forming molecules are amphiphiles, with a hydrophilic "head" and a hydrophobic "tail" on the same molecule. Although we tend to think of membrane lipids as being limited to phospholipids and cholesterol, in fact a surprising variety of amphiphiles take part in membrane structure. Earlier studies (Hargreaves and Deamer, 1978) showed that even single chain amphiphiles such as alkyl phosphates, alkyl sulfates and fatty acids can assemble into bilayer membranes if they contain 10 or more carbons in their hydrocarbon chains.
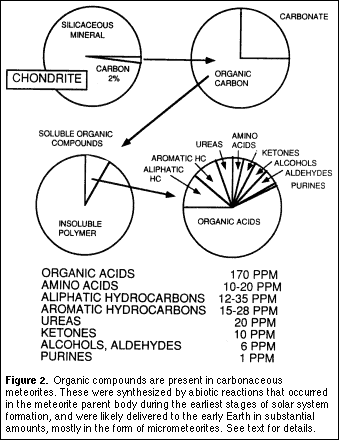
If amphiphilic molecules were present in the mixture of organic compounds available on the early Earth, it is not difficult to imagine that their self-assembly into molecular aggregates was a common process. Is this a plausible premise? A reasonable start is to assume that the mixture of organic compounds in carbonaceous meteorites resembles components available on the early Earth through extraterrestrial infall. Most meteorites are composed of silicon-based minerals, and a small fraction (~5%) of these stony meteorites contain up to several percent of their mass in the form of organic carbon. These are referred to as carbonaceous meteorites, and their organic compounds are actual samples of the chemical components of the early solar system. A kerogen-like insoluble polymer composed largely of covalently linked polycyclic aromatic hydrocarbons is the most abundant organic material, while a series of organic acids (including 10-20 ppm of amino acids) represents the most abundant water-soluble fraction (Figure 1). Aliphatic and aromatic hydrocarbons, ureas, ketones, alcohols, aldehydes and purines are also present.
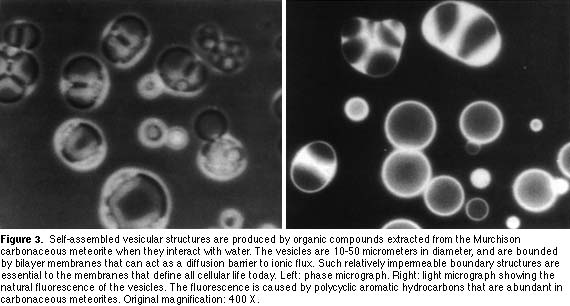
A model protocell
The fact that amphiphilic compounds present in meteorites can self-assemble into bilayer membranes makes it plausible that lipid-like molecules were available on the early Earth and could provide the first cellular boundary structures. The next step is to develop laboratory models of simple cellular systems in which macromolecules are encapsulated by lipid bilayers.
Liposomes are self-assembled spherical lipid bilayers in the size range of bacteria, and provide a useful model system for studies relating to the origin of cellular life. Liposomes are able to capture large molecules such as enzymes and nucleic acids, but their bilayers are relatively impermeable to smaller polar and ionic solutes. In contemporary cells, growth and reproduction require the transport of nutrients across the cell membrane and employ complex protein assemblies to facilitate the transport process. Before such proteins had evolved, what mechanism was available to transport the nutrients required for cell growth?
We have found that bilayer permeability is strongly dependent on chain length. That is, shortening the chains of a given membrane lipid dramatically increases permeation rates of ionic solutes (Paula et al., 1996). We therefore prepared liposomes with lipids of intermediate chain length, using dimyristoyl phosphatidylcholine containing 14 carbon chains. These liposomes can efficiently encapsulate enzymes, yet are permeable enough to allow influx of an externally added substrate molecules.
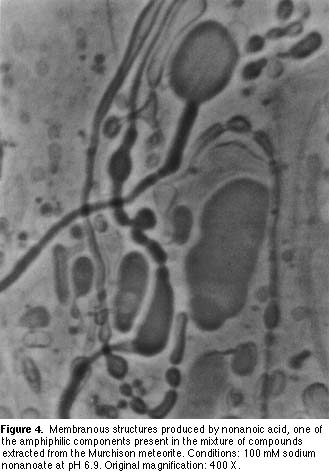
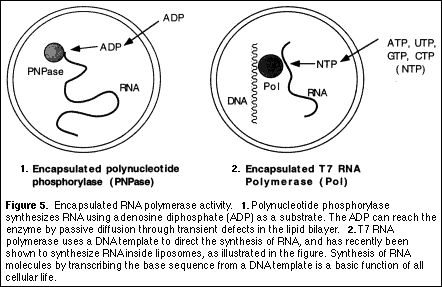
Can life be made in the laboratory? Future directions
These principles can be applied to propose a laboratory version of a protocell. Such a molecular system should have the following properties:
1. A polymer such as a nucleic acid must be replicated by a template-directed polymerization process. The polymer must be able to contain one or more sequences of monomers that correspond to genetic information.
2. A catalytic activity must be present that is somehow linked to the replication process, so that variations in replication affect the rate or efficiency of the catalyzed reaction. The catalyst must also be reproduced by the polymerization process, using the genetic information in the polymer.
3. The replication and catalytic system must be compartmentalized within a membrane-bounded volume so that selection of variations can lead to "speciation" of the encapsulated genetic material.
4. The boundary membrane itself must be able to grow. This could be accomplished either by accumulation of amphiphiles from the environment or by conversion of precursor molecules into amphiphiles. Furthermore, the growth must somehow be coupled to the replication process so that it neither lags behind nor gets too far ahead of the RNA production.
5. There must be a mechanism that allows the assembly to break up into smaller structures when it reaches a given size, and the smaller structures in turn should incorporate the capabilities of the larger system.
Is there any hope that such an assemblage of molecules might be established in the near future? We can begin by considering ribozymes, which are strands of RNA with catalytic sites resembling those of enzymes. Because ribozymes have the potential to act both as catalysts and carriers of genetic information, they have been proposed as the primeval genetic material (Joyce et al., 1987). In one laboratory model of a replicating RNA system, a reverse transcriptase first produces a DNA strand by using a specific ribozyme as a template, and a second DNA polymerase makes multiple copies from the DNA, thereby amplifying the original RNA strand thousands of times as the cycle is repeated. Significantly, the ribozyme itself can also evolve under these conditions when a suitable selective pressure is present. For example, Beaudry and Joyce (1992) found that it was possible to produce a specific catalytic site on a ribozyme by continuously selecting for that site with biochemical hurdles, a kind of molecular breeding carried out in the test tube. Wilson and Szostak (1994) went on to show that a specific catalytic site could be selected from a mixture of trillions of random RNA sequences, similar to the kind of selection that would have occurred in an population of early molecules competing for a resource.
Could such ribozyme systems be considered to be alive in some sense? After all, the RNA is capable of growth and evolution, two primary properties of the living state. However, the system falls short of the definition of life for at least two reasons. First, the ribozyme amplification requires protein enzymes that do not themselves replicate. And second, the test tube acts as a macroscopic encapsulated environment that maintains the system components in contact with one another while the investigator adds monomers and energy. If the components were released from the bounds of the test tube environment, they could not function.
But what if we encapsulated ribozymes in liposomes? It is not at all difficult to set up a system of encapsulated ribozymes so that each liposome contains a different RNA sequence, each with the potential to exhibit a different catalytic activity. Then we might be able to select for a ribozyme that had polymerase activity, because only that liposome would contain RNA, even though no protein enzyme was present. We might also select for a ribozyme that could catalyze the synthesis of phospholipid from activated intermediates such as acyl coenzyme A and a lysophosphatide. Again, only that liposome would increase in volume, because it would be able to synthesize membrane components from non-membranous precursor molecules. None of the proposed ribozyme catalysts have yet been discovered, although a polymerase-like activity in certain ribozymes has been observed (Been and Cech, 1988). Furthermore, virtually nothing is known about the interactions of ribozymes in catalyzing a lipid synthesis reaction. Nonetheless, this thought experiment shows that it is at least possible to conceive of a cellular life form with RNA catalysts that has the potential to be modeled under laboratory conditions. Further investigations of encapsulated replicating catalytic systems will help us to better understand what happened over 3.5 billion years ago as self-assembled molecular systems first began to grow, reproduce, and evolve toward the earliest forms of microbial life.
Acknowledgments
Portions of this chapter were adapted from reviews published elsewhere (Deamer, 1998, 1999).
References cited
Anders, E. 1989. Pre-biotic organic matter from comets and asteroids. Nature 342:255-257.
Bada, J.L., C. Bigham, and S.L. Miller. 1994. Impact melting of frozen oceans on the early Earth — Implications for the origin of life. Proc. Natl. Acad. Sci. USA 91:1248.
Baross, J.A., and S.E. Hoffman. 1995. Submarine hydrothermal vents and associated gradient environments as sites for the origin and evolution of life. Origins of Life 15:327.
Beaudry, A.A., and G.F. Joyce. 1992. Directed evolution of an RNA enzyme. Science 342:255-257.
Been, M.D., and T.R. Cech. 1988. Science 239:1412.
Chakrabarti, A., R.R. Breaker, G.F. Joyce, and D.W. Deamer. 1994. Production of RNA by a polymerase protein encapsulated within phospholipid vesicles. J. Mol. Evol. 39:555-559.
Chyba, C.F., and C. Sagan. 1992. Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: An inventory for the origin of life. Nature 355:125-13.
Corliss, J.B., J.A. Baross, and S.E. Hoffman. 1981. An hypothesis concerning the relationship between submarine hot springs and the origin of life on Earth. Oceanol. p 59. Acta. Proc. 26th Intl. Geolog. Congress, Geology of the Oceans symposium, Paris.
Cronin, J.R., S. Pizzarello, and D.P. Cruickshank. 1988. Organic matter in carbonaceous chondrites, planetary satellites, asteroids and comets. p. 819-857. In J.F. Kerridge and M.S. Matthews (eds.), Meteorites and the Early Solar System. University of Arizona Press, Tucson.
Deamer, D.W. 1999. The first living organisms: In the light or in the dark? ChemTracts (in press).
Deamer, D.W. 1998. Membrane compartments in prebiotic evolution. p. 189-205. In A. Brack (ed.), The Molecular Origins of Life. Cambridge University Press, Cambridge UK.
Delsemme, A. 1984. The cometary connection with prebiotic chemistry. Origins of Life 14:51-60.
Ferris, J.P., and W.J. Hagan. 1984. HCN and chemical evolution: The possible role of cyano compounds in prebiotic synthesis. Tetrahedron 40:1093.
Fox, S. W., and K. Harada. 1958. Thermal copolymerization of amino acids to a product resembling protein. Science 128:1214.
Gavino, V., and D.W. Deamer. 1982. Purification of acyl CoA: 1-acyl-sn-glycerophosphorylcholine acyltransferase. J. Bioenerg. Biomembr. 14:513-526.
Goldacre, R.J. 1958. Surface films: Their collapse on compression, the shapes and sizes of cells, and the origin of life. p. 12-27. In J.F. Danielli, K.G.A. Pankhurst, and A.C. Riddiford (eds.), Surface Phenomena in Biology and Chemistry. Pergamon Press, New York.
Haldane, J.B.S. 1929. The Origin of Life. The Rationalist Annual 148:3-10.
Hargreaves, W.R., and D.W. Deamer. 1978. Liposomes from ionic, single-chain amphiphiles. Biochemistry 17:3759-3768.
Hargreaves, W.R., S. Mulvihill, and D.W. Deamer. 1977. Synthesis of phospholipids and membranes in prebiotic conditions. Nature 266:78-80.
Holland, H.D. 1984. The Chemical Evolution of the Atmosphere and Oceans. Princeton University Press, Princeton.
Joyce, G. F., A.W. Schwartz, S.L. Miller, and L.E. Orgel. 1987. The case for an ancestral genetic system involving simple analogues of the nucleotides. Proc. Natl. Acad. Sci. USA 84:4398.
Kasting, J., and T.F. Ackerman. 1986. Climatic consequences of very high carbon dioxide levels in the Earth's early atmosphere. Science 234:1383.
Kvenvolden, K.A., J.G. Lawless, K. Pering, E. Peterson, J. Flores, C. Ponnamperuma, I.R. Kaplan and C. Moore. 1970. Evidence for extraterrestrial amino acids and hydrocarbons in the Murchison meteorite. Nature 28:923.
Miller, S.L. 1953. Production of amino acids under possible primitive Earth conditions. Science 117:528.
Miller, S.L. and H.C. Urey. 1959. Organic compounds synthesized on the primitive Earth. Science 130:245.
Mojzsis, S.J., G. Arrhenius, K.D. McKeegan, and T.M. Harrison. 1996. Nature 384:55.
Monnard, P.-A., W. Vercoutere, and D.W. Deamer. (unpublished results).
Oro, J. 1961. Comets and the formation of biochemical compounds on the primitive Earth. Nature 190:389-390.
Oro, J., E. Sherwood, J. Eichberg, and D. Epps. 1978. Formation of phospholipids under primitive Earth conditions and the role of membranes in prebiological evolution. p. 1-21. In D.W. Deamer (ed.), Light-Transducing Membranes: Structure, Function and Evolution. Academic Press, New York.
Pace, N.R. 1991. Origin of life — Facing up to the physical setting. Cell 65:531.
Pace, N.R. 1997. Science 276:734.
Paula, S., A.G. Volkov, A.N. Van Hoek, T.H. Haines, and D.W. Deamer. 1996. Permeation of protons, potassium ions and small polar molecules through phospholipid bilayers as a function of membrane thickness. Biophys. J. 70:339-348.
Schopf, J.W., and B.M. Packer. 1987. Early Archean (3.3-billion to 3.5-billion-year-old) microfossils from Warrawoona Group, Australia. Science 237:70.
Stevens, T.O., and J.P. McKinley. 1995. Science 270:450.
Usher, D. 1977. Early chemical evolution of nucleic acids: A theoretical model. Science 196:311-313.
Wilson, C., and J.W. Szostak. 1994. In vitro evolution of a self-alkylating ribozyme. Nature 374: 777.
Woese, C.R. 1987. Bacterial evolution. Microbiol. Rev. 51:221.
Wright, M.C., and G.F. Joyce. Continuous in vitro evolution of catalytic function. 1997. Science 274:1309.