Archosauria
The "ruling reptiles"
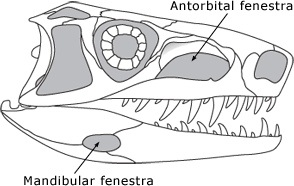 The archosaur skull includes two new fenestra (skull openings). The first of these is the mandibular fenestra, an opening through the jawbone. The other is the antorbital fenestra, a term that means "the opening in front of the eye." |
The ancestral archosaurs probably originated some 250 million years or so ago, in the late Permian Period. Their descendants (such as the dinosaurs) dominated the realm of the terrestrial vertebrates for a majority of the Mesozoic Era. Today, only the birds and crocodilians exist to provide a glimpse into the past glory of archosaurs.
Systematics
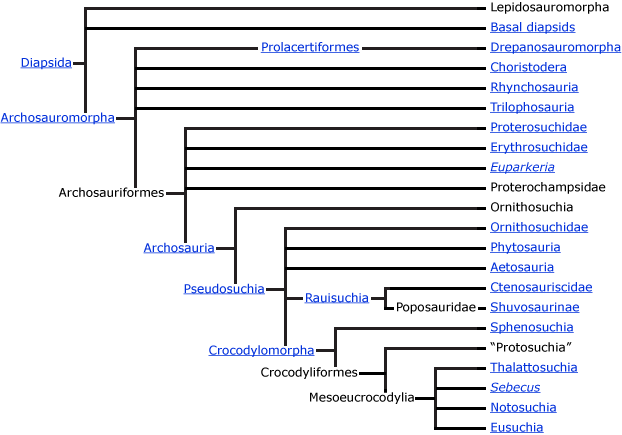
Archosauria is defined as the group that includes the common ancestor of crocodiles and birds and all of its descendants. Because the group is defined this way, all archosaurs fall into one of two big clades: Ornithosuchia, or archosaurs on the evolutionary line to birds, and Pseudosuchia, or archosaurs on the evolutionary line to crocs. Ornithosuchia includes dinosaurs, dinosaur relatives such as Marasuchus, pterosaurs, and a few oddballs like Scleromochlus, whose relationships are a bit uncertain. In addition to crocs, Pseudosuchia includes the croc-like phytosaurs, armadillo-like aetosaurs, carnivorous ornithosuchids, and rauisuchians, which themselves included both large carnivores and toothless, beaked animals that probably ate plants.

Morphology
The archosaurs are characterized by numerous synapomorphies that lend strong support to the hypothesis that they form a monophyletic group (clade) exclusive of other Reptilia. First of all, the "stem archosaurs" (properly termed Archosauromorpha), including Champsosauridae and Euparkeria, have a calcaneal tuber. This is a bony process projecting posteriorly from the ankle joint that serves as an attachment point for some of the lower leg flexor muscles. If you feel your heel, the bone that forms it is your calcaneal tuber. This is an example of convergent evolution; synapsids and archosaurs evolved these features independently.
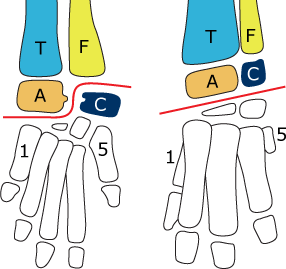 The two major groups of archosaurs are distinguished by differences in the joint of their ankle. The crocodiles and their relatives (Pseudosuchia) have a crurotarsal ankle joint (left), while dinosaurs and pterosaurs (Ornithosuchia) have a mesotarsal ankle joint (right). The red line in each image shows the plane of the ankle hinge. (T=tibia, F=fibula, A=astragalus, C=calcaneum) |
True archosaurs (the Archosauria proper), including the major stem groups Pseudosuchia (the crocodilians and their relatives) and the Ornithosuchia (the birds and their relatives) evolved yet more new features. The fifth toe in the foot, homologous with your "pinky toe" was reduced in size. In earlier vertebrates, the palate (roof of the mouth) bore at least one row of accessory teeth, but the archosaurs appear to have lost this feature, as did many other lineages of tetrapods. Finally, the two main lineages of archosaurs each have a specialized ankle joint that changed differently in each group, as shown below.
The Pseudosuchia have what is called a crurotarsal, or "crocodile-normal" ankle. This is a very flexible arrangement in which the astragalus (medial proximal ankle bone) bears a peg that fits into a socket in the calcaneum (lateral proximal ankle bone). Ankle rotation then occurs between these two bones, permitting both a somewhat erect stance (like the crocodilian "high walk") where the hindlimb is held closer to the midline of the body, and a more sprawling stance like that of earlier tetrapods. The result of this is that the pseudosuchians can move in two different ways; walking with an erect or sprawling posture.
The Ornithosuchia have a mesotarsal ankle, which is a simple hinge joint between the lower leg and astragalus and calcaneum, and the distal ankle bones. This restricts the posture to a more erect orientation, so the gait can be called parasagittal — the limbs move parallel to the vertebral column, and are held relatively vertical. Birds and most mammals have this parasagittal gait; birds inherited it from their dinosaurian ancestors, while mammals evolved it independently. The advantage of a parasagittal gait might be that it improves maneuverability/agility. A concordant disadvantage would be that it reduces stability. It's easier to tip over an erect cow than it is to tip over a more sprawling crocodile of similar size; the crocodile has a wider base of support, and thus can be said to be more stable. However, the relationship between stance and stability is still not completely understood, so it's difficult to say exactly what advantages an erect stance has. Another proposed advantage of an erect stance and parasagittal gait was that it was more efficient, but this has not held up to experimental analysis — erect animals move about as efficiently as similarly-sized sprawling ones.
It is difficult to say what sorts of soft tissues extinct archosaurs really had, but the fact that both crocodilians and birds have a four-chambered heart lends support to the notion that this is a trait inherited from a common ancestor (and another convergence with mammals). Birds and crocodiles also share expanded pneumatic sinuses in their skulls — and elsewhere in bird skeletons — which can also be seen in fossil archosaurs, such as the hadrosaurs, so the expansion of these craniofacial air sinuses is another trait shared by archosaurs. The sinuses appear to serve as skull-remodeling agents, reshaping the bones of the skull in response to stresses and other influences.
The fossil record
Students of archosaur evolution are blessed with a wonderful fossil record for many groups of archosaurs, including some very bizarre extinct taxa. The first archosauromorphs (relatives of the true archosaurs) appear in the fossil record in the Early Triassic, about 245 million years ago, just after the great end-Permian extinction. They include weird hippo-size beaked herbivores called rhynchosaurs, long-necked reptiles called prolacertiforms, evil-looking terrestrial predators like the erythrosuchids and proterosuchids, and close relatives of the true archosaurs, including Euparkeria. Many of these early groups are limited to the Triassic Period, not enduring the extinctions in the Upper Triassic that the dinosaurs and other taxa survived.
The Upper Triassic (about 225 million years ago) witnessed the appearance of several new groups of archosaurs, some of which have living descendants today. Most other archosaurs and archosauromorphs did not make it into the Jurassic, but the crocodilian lineage did, as did the dinosaurs and the tenacious champsosaurs, which all appeared at about the same time in the Upper Triassic. Close relatives of the dinosaurs such as the pterosaurs and the possibly dinosaurian Eoraptor and Herrerasaurus also show up in the Upper Triassic.
The survivors of the Upper Triassic-Lower Jurassic extinctions went on to dominate the rest of the Mesozoic Era, but not without hitches. The Upper Cretaceous Period (around 70 million years ago) was a time of global change and "ecological reshuffling," when many taxa did not make the cut. Pterosaurs, many dinosaurs (including some major bird groups), and some crocodilians vanished around this time.
As the Cenozoic Era dawned 65 million years ago, things were different — crocodiles and champsosaurs were still doing well, but the dinosaurs had taken a one-two punch, and only the neornithine birds persevered to continue the massive diversification that began in the Cretaceous Period. Today we are left with only the crocodilians and the birds as extant (living) archosaurs.
Something to keep in mind
The important thing to remember about archosaurs is that the living forms, birds and crocs, are not typical representatives of their respective groups! Many ornithosuchians, like Pteranodon, Apatosaurus, and Triceratops, were not particularly bird-like, although they share important characters with birds. Similarly, most pseudosuchians lived on land, and quite a few ate plants, unlike living crocs. In fact, the direct ancestors of living crocodiles were long-legged running animals the size of chihuahuas that ate insects. So when you think of Archosauria, remember that it includes animals that walked on two legs and on four, fliers and swimmers, tiny hummingbirds the size of your thumb and giant sauropods the size of a house; animals with saber-like teeth and others with toothless beaks, that ate everything from nectar to pine cones and ants to antelopes — and that only a handful of these animals were semi-aquatic fish-eaters like alligators.
Text by John R. Hutchison, 10/1995, with additions by Brian R. Speer and John, 1/1998; additional text by Matt Wedel, 5/2007.