The Rudists
Rudists lived in shallow marine environments from the Late Jurassic to the Late Cretaceous periods. They had two asymmetric valves with one valve attached to the sea floor. Today, their fossils are found throughout the tropics in the Mediterranean, the Middle East, the Caribbean, and Southeast Asia. Rudists dominated the world of reefs throughout the Cretaceous until the Cretaceous-Tertiary mass extinction event that caused their demise.
Fossil record
 Rudist fossils in natural position. |
Preservation
The hard outer shells of the Rudists were recrystallized from aragonitic or calcitic shells to calcitic fossils. Preservation bias needs to be considered when studying the fossil record of rudists. Shells made out of aragonite are less likely to be preserved than shells made out of calcite because of there chemical properties. What this means, for example, is that Radiolitidae (a calcitic shelled rudist) may be overrepresented in the fossil record when compared to Caprinidae (an aragonitic shelled rudist) (Steuber and Löser 2000).
Biogeography
Rudist reefs were located in the equatorial latitudes, generally around what are now Meso-America, the Mediterranean, Northern Africa, the Middle East, and Southeast Asia. Rudists were especially abundant during the periodic occurrence of an extra salty, extra warm seaway described as the Supertethys (Arthur et al. 1996). Local and global climate changes have been inferred by studying the biogeographic extent of rudist dominated ecosystems at certain times. For instance, the constriction of rudist reefs towards the equator during the middle Cretaceous may have been caused by the movement of heated waters away from the tropical zone (Arthur et al. 1996). In this case, the Supertethyan circumstances were thought to have collapsed, which in turn, led to rudist reef collapse.
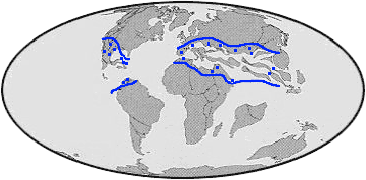 Extent of rudist reefs during the Cretaceous. |
 |
Life history & ecology
Rudists grew by accretion (or the increase in size by addition) and were suspension feeders. Generally, large-bodied rudists lived longer and reproduced relatively frequently with small spawns, while small-bodied rudists lived shorter lives and reproduced with less frequent, but large, spawns (Gili et al. 1995).
Rudists lived in shallow ocean waters on the sea floor. These organisms were epifaunal, which means they were usually attached to the sea floor sediment. The clustering and building up of Rudist habitats caused the creation of "Rudist Reefs" which were the dominant reef frameworks in the Cretaceous oceans.
More on morphology
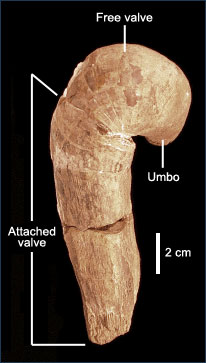
Basic external features of rudists include the umbo and thick, asymmetric right and left valves. The umbo is the rounded protrusion found just above the hinge, and the hinge is the pivoting point where the two valves meet. There are three main valve plans found in rudists. These plans (from Perkins 1969), are based on relative size of the valves and the level of valve coiling:
 Notice the tooth near the center of this valve from Masneronia. |
Important characteristic bivalve features include teeth and sockets which are found in the hinge to prevent shell misplacement, adductor muscles to pull the valves together, and ligamental structures for the movement of the valves. In younger rudist families, the ligament function is lost (which will be discussed in more detail later) and there is a question about what replaced this function. Skelton (1976) believes the shell did not open fully and the small opening that existed between the valves was enough for feeding and waste processes. On the other hand, Seilacher (1998) suggests the existence of diductor muscles for opening the valves, but this has yet to be fully proven. Soft tissues are rarely preserved in the fossil record so the study of rudist organs can be a difficult task.
 |
 |
||
|
At left, a longwise section through Coralliachama orcutti. At right, a cross section of Monopleura salazari. |
|||
Johnson and Kauffman (1988) summarized important evolutionary changes in rudist morphology. They pointed out that earlier rudists tended to have wide, and more coiled, bases, while later rudists had thinner bases, more erect forms, and more ornamentation. Sea floor attachment bases became smaller through time while the attached valve went from coiled to more cone-like forms. In later rudist types the free valve became smaller and less coiled.
One important innovation in rudist morphology was the ligamental groove, or invagination of the ligament, which first became distinctive in Caprotinidae (Perkins 1969). This ligamental groove allowed for more uncoiled shell designs (Steuber 1999). Finally, in Radiolitidae and Hippuritidae, the loss of the functional ligament allowed for an upright growth (Steuber 1999, Seilacher 1998).
Systematics
There are 792 described and named species of rudists found in the central-eastern Mediterranean and Middle East (Barremian-Maastrichtian) (Steuber 1999) and 214 described species of rudists found in the Carribean province (Johnson and Kauffman 1990). By looking at their hinge structure, bottom attachment, and several internal features, rudists have been classified according to the families in the table below.
|
Characteristics of rudists include the following (from Dechaseaux):
Rudists were derived from megalodontids (Hippuritoida-Megalodontia), a bivalve group that shared similar morphological states with the earliest rudists (Skelton and Smith 2000). No direct descendants of rudists exist today as the abrupt mass extinction wiped out the entire group during the Maastrichtian. See Steuber's (1999) tentative phylogeny below right.
Sources
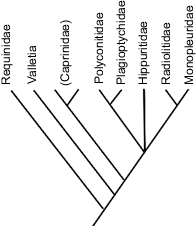 A tentative phylogeny of Hippuritoida (the order rudists belong to) by Steuber 1999. |
Original text and photos of Coralliachama orcutti, Monopleura salazari, and Masneronia by Cherry Zamora, 2002. Photo of rudist fossils in place by Carole Hickman, UCMP.